


Malnutrition in MASLD-related hepatocellular carcinoma: assessment limitations, multidimensional mechanisms, and microbiome-based interventions
 0
0 Abstract
With the rapid rise in hepatocellular carcinoma (HCC) incidence associated with metabolic dysfunction-associated steatotic liver disease (MASLD), malnutrition has emerged as a critical comorbidity that affects treatment tolerance and survival outcomes. This review systematically examines the challenges of assessing malnutrition in metabolic dysfunction-associated steatotic liver disease-related hepatocellular carcinoma (MASLD-HCC), its multidimensional pathological mechanisms, and emerging intervention strategies. In current clinical practice, the absence of specialized assessment tools limits the effectiveness of existing evaluation systems in addressing distinct phenotypes, such as obesity and ascites. The underlying mechanisms are complex; disruptions of the “gut-liver-muscle axis”, centered on intestinal dysbiosis, form a key network driving metabolic reprogramming, chronic inflammation, and muscle wasting. Regarding interventions, microbiome-targeted therapies - including fecal microbiota transplantation and probiotics/synbiotics - have shown promise in preclinical studies for modulating immunometabolism and improving nutritional status. However, their translation into routine clinical practice remains constrained by major bottlenecks, including insufficient large-scale randomized controlled trial evidence, uncertainty regarding long-term safety, and inadequate standardization. Future efforts should urgently focus on establishing precise assessment systems and conducting high-quality clinical validation.
Keywords
INTRODUCTION
Metabolic dysfunction-associated steatotic liver disease (MASLD) has emerged as one of the leading etiological factors for hepatocellular carcinoma (HCC) worldwide. Based on data from the past three decades, the contribution of traditional dominant factors (viral hepatitis and alcohol) has gradually declined, whereas the proportion of cases attributable to MASLD has increased annually by 0.66% (95%CI: 0.64%-0.67%)[1], consistent with previous findings[2,3].
Malnutrition is a common complication of HCC, with a reported prevalence of approximately 25% among HCC patients[4,5]. It is directly associated with unfavorable treatment outcomes, reduced quality of life, and increased mortality in HCC[6,7]. The mechanisms contributing to malnutrition in MASLD-related HCC may include insulin resistance (IR), metabolic energy reprogramming, chronic inflammation, gut-liver axis dysregulation, and anorexia-associated behavioral changes[8-10].
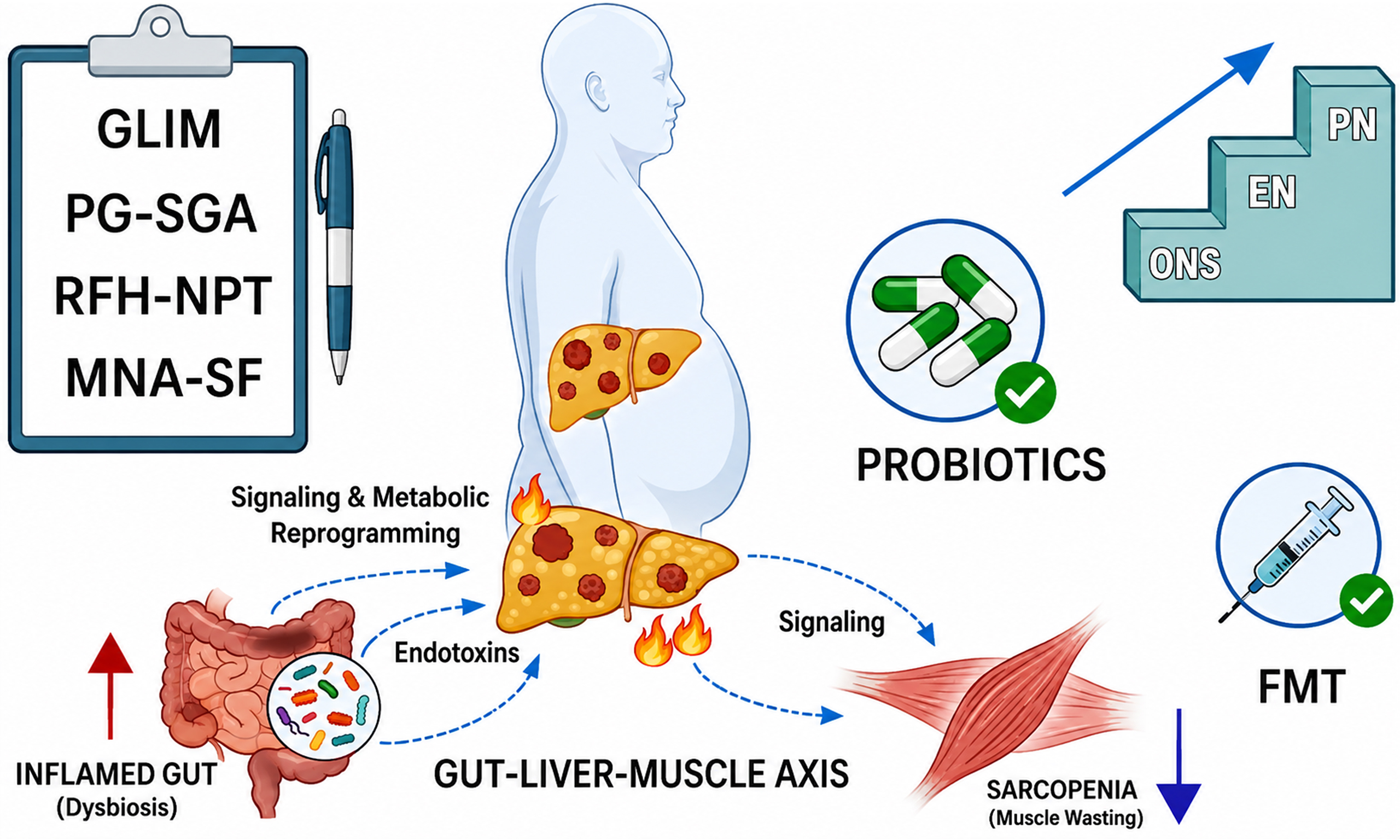
At present, there is a lack of screening and assessment tools specifically designed for metabolic dysfunction-associated steatotic liver disease-related hepatocellular carcinoma (MASLD-HCC) in routine clinical practice, and general tools such as Nutritional Risk Screening 2002 (NRS-2002) and the Patient-Generated Subjective Global Assessment (PG-SGA) remain widely used[11]. However, these tools have limitations in accurately evaluating specific indicators, including obesity phenotype, fluid retention, and hypoalbuminemia, which results in a high risk of underdiagnosis. To improve assessment accuracy, clinical practice may consider combined strategies, such as incorporating the Royal Free Hospital-Nutritional Prioritizing Tool (RFH-NPT), integrating imaging examinations or anthropometric measurements, and including inflammatory biomarkers[12-14].
Current nutritional support strategies primarily focus on improving clinical indicators and alleviating patient discomfort[15]. Microbiota-targeted therapies, such as probiotics and fecal microbiota transplantation (FMT), may provide new perspectives, although large-scale clinical validation is still lacking. This article offers a concise overview of existing nutritional assessment tools, pathogenic mechanisms, and intervention strategies for malnutrition in MASLD-HCC, with the goal of informing clinical practice and guiding future research.
Currently, there are no specific nutritional screening and assessment tools designed for patients with MASLD-HCC in routine clinical practice. For malnutrition risk screening in adult oncology outpatients, the American Society for Parenteral and Enteral Nutrition (ASPEN) has classified tools into different levels according to usability: the Malnutrition Screening Tool (MST) is classified as Level 1, the PG-SGA as Level 2, and the Malnutrition Universal Screening Tool (MUST) and the NRS-2002 as Level 3[16]. However, Ge et al. reported that although MST is the fastest, its sensitivity is too low (28.5%), and it is recommended only for community settings[17]. The scored Patient-Generated Subjective Global Assessment (sPG-SGA) has been established as a standard nutritional assessment method for cancer patients and is widely used in clinical practice[18]. In a retrospective analysis of 3,777 adult hospitalized cancer patients, the GLIM criteria showed moderate agreement with sPG-SGA[19]. Another study found that, without initial screening using NRS-2002, GLIM demonstrated greater concordance with PG-SGA for diagnosing malnutrition[20]. In addition, for elderly cancer patients, the Mini Nutritional Assessment Short Form (MNA-SF) has also shown good applicability[21].
However, none of the tools above are specifically designed for HCC patients. HCC is frequently accompanied by sodium and water retention and hypoalbuminemia; ascites may therefore mask weight loss related to muscle or fat loss, potentially compromising the accurate interpretation of existing tools. In contrast, screening tools designed specifically for malnutrition in liver cirrhosis partly address this limitation. Clinical studies have shown that the Liver Disease Undernutrition Screening Tool (LDUST) appears to be more accurate than MUST and NRS-2002 in assessing nutritional status in patients with ascites[22]. The 2019 European Society for Clinical Nutrition and Metabolism (ESPEN) guidelines also recommend using the Royal Free Hospital Nutritional Prioritizing Tool (RFH-NPT) for nutritional screening[15]. Although RFH-NPT has drawbacks - such as a longer administration time and the requirement for a professional dietitian to be involved[23] - other studies have indicated that RFH-NPT and LDUST are the only screening tools proven to accurately detect malnutrition in patients with cirrhosis[24].
With a focus on MASLD-HCC, one of its important features is sarcopenic obesity[25]. Assessment tools that rely on low body mass index (BMI) for diagnosis may be biased. A study using the Controlling Nutritional Status (CONUT) index as the assessment standard found no significant difference in the prevalence of malnutrition between obese (BMI ≥ 30) and non-obese MASLD patients[25]. In terms of body composition assessment, the skeletal muscle index at the third lumbar vertebra measured by CT is considered the gold standard, whereas the psoas muscle thickness-to-height ratio (PMTH) offers advantages in simplicity and speed[26].
In summary, there is currently no specific screening and assessment tool for nutritional status tailored to a single type of cancer in routine clinical practice. Combining different tools is therefore necessary to improve diagnostic accuracy. For example, combining RFH-NPT with mid-upper arm circumference (MUAC) or mid-upper arm muscle circumference (MUMC) can significantly improve the diagnosis of sarcopenia[27]. Artificial intelligence is promising for integrating multimodal data and enabling dynamic prediction, but it currently lacks support from large-scale datasets[28]. For details, see Table 1.
Different screening and assessment tools
| Screening tool | Reference standard | Sample source | Assessment dimensions | Sensitivity | Specificity | AUC | Ref. | Advantages | Limitations |
| GLIM | sPG-SGA | 3,777 adult cancer inpatients | 1. At least one phenotypic criterion and one etiologic criterion should be combined | 70.5% | 88.3% | 0.79 | [19] | Internationally unified, dual-dimensional (phenotypic + etiologic), compatible with existing screening tools, and highlights sarcopenia | Relatively complex, and some items may be influenced by subjective factors |
| PG-SGA | GLIM | 637 adult cancer patients | 1. The patient-completed component includes, specifically, weight loss, nutrition-impact symptoms, dietary intake, and functional capacity 2. The clinician-completed component assesses disease and age, metabolic stress, and physical examination | 81.1% | 71.6% | 0.763 | [29] | Preferred in oncology settings, with good predictive performance | Time-consuming and requires trained personnel |
| NRS-2002 | GLIM | 637 adult cancer patients | 1. BMI 2. Dietary intake 3. Disease severity | 81.7% | 97.6% | 0.896 | [29] | Content and predictive validity: moderately reliable; practical; considers disease severity | Weight from fluid collections (ascites, peripheral edema) is not accounted for |
| MUST | GLIM | 1. BMI 2. Unintentional weight loss and acute disease compromising nutritional intake for more than 5 days | 52.2% | 99.1% | 0.757 | Validated in both inpatients and outpatients; rapid and simple | Weight from fluid collections (ascites, peripheral edema) is not accounted for, and disease severity is not considered | ||
| MNA-SF | GLIM | Forty cancer patients (mean age 84.8 ± 5.5 years) | 1. BMI 2. Recent weight loss 3. Appetite changes 4. Mobility 5. Psychological stress and neuropsychological problems | 100% | 50% | 0.75 | [21] | Simple, non-invasive, and economical | Primarily targeted at elderly patients |
| AIWW | PG-SGA | 11,360 cancer patients | 1. Age 2. Dietary intake 3. Weight loss 4. Walking ability | 93.40% | 46.10% | 0.785 | [17] | Very high sensitivity with rapid completion | Low specificity; requires validation with GLIM |
| RFH-NPT | GLIM | 78 patients with liver cirrhosis | 1. Alcoholic hepatitis or tube feeding 2. Fluid overload and its impact on food intake and weight loss 3. BMI, unplanned weight loss, and daily dietary intake | 90.91% | 60.00% | 0.755 (0.646-0.863) | [30] | A liver disease-specific tool; rapid and simple; reduces the impact of fluid retention | Requires further validation and broader recognition |
| SGA | 80 patients with liver cirrhosis | 97.3% (90.7, 99.7) | 74.4 % (57.9, 87.0) | 0.95 | [31] | ||||
| LDUST | RFH-NPT | 207 patients with liver cirrhosis | 1. Food intake 2. Weight loss 3. Fat and muscle depletion 4. Fluid accumulation 5. Functional capacity | 92.1% | 67.2% | 0.797 | [32] | Includes a fluid retention item, is rapid and self-administered, and has high sensitivity | Relies entirely on patients’ subjective judgment and has a low negative predictive value |
MULTIDIMENSIONAL MECHANISMS UNDERLYING MALNUTRITION ASSOCIATED WITH HCC
Gut-liver-muscle axis
Obesity, IR, MASLD, and sarcopenia form a dynamic network that promotes the progression from metabolic dysfunction to malignant transformation through the gut-liver-muscle axis. In the early stage of disease, visceral fat accumulation drives IR via the release of free fatty acids (FFAs). IR, on the one hand, enhances
Notably, as the largest peripheral glucose reservoir, the progressive loss of skeletal muscle further undermines systemic insulin sensitivity, establishing a self-reinforcing loop of “IR-sarcopenia-MASLD”. Large-scale cohort studies have confirmed that this association is independent of obesity[34,35]. Within this pathological cascade, gut microbiota dysbiosis acts as a central hub linking MASLD to HCC-associated malnutrition. Clinical evidence indicates that HCC patients display distinct microbial imbalances superimposed on a MASLD background: a pronounced depletion of beneficial butyrate-producing taxa such as Faecalibacterium, alongside enrichment of potentially pathogenic genera including Streptococcus and Veillonella. In addition, small intestinal bacterial overgrowth (SIBO) is highly prevalent in MASLD populations, reaching up to 35%, and is frequently accompanied by diarrhea and vitamin B12 deficiency[36]. This dysbiosis disrupts intestinal barrier function through multiple molecular mechanisms: tumor necrosis factor-alpha (TNF-α) promotes the internalization of occludin, thereby impairing tight junctions; interleukin-13 (IL-13) upregulates claudin-2, creating a “pore pathway”; and zonulin, a key permeability regulator, further aggravates “leaky gut” via the PAR2-EGFR axis[37,38]. Compromised barrier integrity facilitates the translocation of lipopolysaccharide (LPS) and viable bacteria to the liver[39], where they activate innate immunity through the Toll-Like receptor 4 (TLR4)/nuclear factor-kappa B (NF-κB)/c-Jun N-terminal kinase (JNK) signaling pathway[40], thereby accelerating the progression of MASLD to HCC[41,42]. Critically, microbiota-immune-muscle crosstalk shapes the phenotype of cachexia. The chronic inflammatory milieu induced by LPS translocation directly drives muscle wasting: circulating levels of TNF-α and IL-6 are markedly elevated in MASLD-HCC patients with sarcopenia[43-45]. Mechanistically, TNF-α promotes muscle atrophy by accelerating ubiquitin-proteasome-mediated protein degradation, whereas inhibition of the IL-6/STAT3 pathway has been shown to enhance muscle repair in HCC mouse models[46-50]. Meanwhile, microbial metabolic reprogramming is pivotal. Short-chain fatty acid (SCFA) deficiency not only impairs GPR41/43-mTOR-mediated anabolic signaling in muscle but also suppresses protein translation by competitively consuming branched-chain amino acids (BCAAs)[51,52]. Moreover, reduced SCFAs are accompanied by lower glucagon-like peptide-1 (GLP-1) levels, further worsening IR and forming a vicious cycle of “metabolic dysfunction-sarcopenia”[53]. Genetic factors also modulate this network; for example, the patatin-like phospholipase domain-containing protein 3 (PNPLA3) rs738409 G allele exacerbates liver injury in the setting of low muscle mass[54].
In summary, the gut microbiota forms the core pathological network of MASLD-HCC-associated malnutrition by mediating barrier disruption, inflammatory cascades, and metabolite imbalances. Microbiota- or metabolite-targeted intervention strategies hold significant translational potential[55].
Metabolic reprogramming
The hypermetabolism of tumor cells is primarily manifested by the Warburg effect and the Cori cycle. The Warburg effect describes the preference of tumor cells for glycolysis to generate ATP, even under aerobic conditions, leading to the release of large amounts of lactate[56]. Excess lactate enters the Cori cycle and is converted back into glucose via hepatic gluconeogenesis for systemic use[57]. This cyclical process results in inefficient energy expenditure[58], further increasing resting energy expenditure. Studies have shown that lactate, as a critical signaling molecule, can directly activate proopiomelanocortin (POMC) neurons and suppress appetite through astrocyte-neuron communication[59]. In addition, Liu et al. demonstrated that lactate induces browning of white adipose tissue via the adipocyte-specific G protein-coupled receptor (GPR81)[60]. As an intermediate product of metabolic reprogramming, lactate further aggravates malnutrition itself.
Anorexia
Appetite is primarily regulated by two key hypothalamic neuronal populations: agouti-related peptide (AgRP) and POMC neurons[61,62]. Evidence indicates that pro-inflammatory cytokines (IL-1β, IL-6, TNF-α)[63-67], as well as lipocalin 2 (LCN2)[68], prostaglandin E2 (PGE2)[69], and insulin-like 3 (INSL3) produced within the tumor microenvironment, together with leptin and Metrnl secreted by adipose tissue[70], can modulate feeding behavior by acting on the hypothalamus. Borner et al. identified that GLP-1 signaling in the nucleus tractus solitarius is a critical mediator of anorexia in rats with HCC[71]. Further studies have shown that silencing the calcitonin gene-related peptide-parabrachial nucleus (CGRP-PBN) pathway in the nucleus tractus solitarius can reverse anorexia[66], suggesting that the nucleus tractus solitarius also serves as a key hub for appetite regulation.
Gut microbiota can influence appetite by modulating secondary bile acids through the farnesoid X receptor and Takeda G protein-coupled receptor 5 (TGR5), thereby inhibiting ghrelin secretion and stimulating neuropeptide Y (NPY)/AgRP neurons via the phospholipase C-Inositol 1,4,5-Trisphosphate (PLC-IP3)/diacylglycerol-protein kinase C (DAG-PKC) pathway[72]. In turn, SCFA regulate the secretion of peptide YY (PYY) and GLP-1[73], which can bind to GLP-1 receptors in the hypothalamus[74]. This suggests that gut microbiota dysbiosis may simultaneously act on central appetite-regulating centers via vagal stimulation and endocrine pathways, leading to decreased appetite[75-77]. Additionally, reduced appetite is associated with treatment-related side effects, such as taste disturbances[78,79] and diarrhea[80].
NUTRITIONAL INTERVENTION
Oral nutrition, enteral nutrition, and parenteral nutrition form a three-tiered nutritional support strategy commonly used in clinical practice. MyPath, a patient-centered assessment and management system developed in the European Union, provides tailored, guideline-based care options[81]. This program is currently undergoing pilot testing and may, in the future, offer novel insights and experience to address the problem of inconsistent implementation of nutritional guidelines in clinical settings. In addition, the gut microbiota has demonstrated therapeutic potential in the management of various diseases, and we specifically focus on its current application and efficacy in HCC.
Fecal microbiota transplantation (FMT)
FMT is considered to have the potential to reshape the gut microbiota, restore microbial function, mitigate systemic inflammation, and reprogram the tumor microenvironment[82-84]. For instance, early-phase clinical trials in patients with refractory melanoma have demonstrated preliminary efficacy and acceptable safety[85], providing initial proof-of-concept for its use in oncology. However, clinical translation faces multiple barriers. First, donor screening is a core prerequisite to ensure safety and efficacy, yet the standards are extremely stringent. Current screening mainly follows modified blood-donor guidelines, using questionnaires and laboratory tests to exclude individuals at risk of infectious diseases, metabolic disorders, malignancies, and other conditions[86]. As a result, the proportion of eligible donors is very low. For example, in a Korean screening program, only 5% of volunteers met the criteria for repeat donation[87]. Similarly, under European standardized protocols, the eligibility rate is approximately 10%, and regular rescreening is required[88]. This substantially limits the scale and accessibility of donor banks while increasing treatment costs. Second, safety data in the specific population of cancer patients remain particularly scarce. Although approaches such as washed microbiota transplantation[89] and encapsulated formulations[90] aim to reduce risks, adverse events such as fever and vomiting may still occur after FMT[91]. Most importantly, for the endpoint of improving cancer-associated malnutrition, there is almost a complete lack of evidence from large-scale randomized controlled trials[92]. The efficacy of FMT may depend heavily on donor-recipient compatibility and complex microbial network effects[93], underscoring the need to develop personalized donor-matching strategies.
Probiotics and prebiotics
The combined use of probiotics and prebiotics, known as synbiotics, has been redefined as a mixture of live microorganisms and selectively utilized substrates by host microorganisms, intended to confer health benefits[94]. The mechanisms by which synbiotics improve malnutrition are relatively well established: specific strains (e.g., Lactobacillus, Streptococcus faecalis) can reduce circulating LPS levels[95], thereby attenuating a key pathway linking gut dysbiosis to systemic inflammation. In MASLD models, probiotics such as Lactobacillus have been shown to improve IR, reduce inflammation, and restore the abundance of beneficial bacteria[96-98].
Moreover, Lactobacillus rhamnosus GG and Escherichia coli Nissle 1917 have been shown in models to enhance intestinal barrier integrity, increase protective mucus secretion, and improve SCFA metabolism[99,100]. However, orally administered probiotics face challenges such as degradation by gastric acid and bile salts, leading to reduced viability and colonization capacity[101,102], which directly affects the stability and reproducibility of their therapeutic effects. Pre-encapsulation strategies for Lactobacillus species, such as alginate hydrogel beads, Cas, and Alg, have been demonstrated to increase bacterial survival rates, as shown in Table 2.
Results of experiments with different encapsulation materials
| Probiotics | Material | Results | Ref. |
| Lactis Bb12 | Poly(vinyl alcohol) | An increase to 1.71 × 108 was observed after 1 week | [103] |
| L. plantarum 550 | Alginate hydrogel beads | After 6 h of digestion, the loss of viable cells was only 1.09 log CFU/g, indicating a substantial improvement | [104] |
| Lactobacillus acidophilus 11073 | WPI-OSA-starch complex | A significant (P < 0.05) survival rate (95.94% ± 1.64%) was observed after spray drying | [105] |
| L. plantarum NCIMB 8826 | Chitosan gel particles | No significant decrease in cell concentration was observed during a 2-h incubation in simulated gastric fluid at pH 2 (P > 0.05) | [106] |
| L. acidophilus CGMCC1.2686 | Alginate and colloidal particles (TPC-stabilized nanoemulsion, NE)/TPC | ALG + NE exhibited the best protective effect (8.93 ± 0.06 log CFU/g) | [107] |
| L. plantarum FZU3013 | Chitosan (Cas) and alginate (Alg) | Compared with alginate beads, it better preserves bacterial structural integrity | [108] |
| Lactobacillus plantarum | ALG-WPI and ALG-PEC-WPI | The survival rates increased by 65.37% and 72.06%, respectively | [109] |
CONCLUSION
This review systematically examines the distinctive clinical challenges posed by malnutrition in patients with MASLD-HCC. The central finding is that conventional etiology-based nutritional assessment tools - whether general or liver-specific - are increasingly insufficient to accurately capture the dynamically evolving nutritional risk in MASLD-HCC, shaped by the “gut-liver-muscle axis”. This pathogenic network includes intricate interactions among gut dysbiosis, metabolic derangements, systemic inflammation, and anorexia-related behaviors, presenting both a barrier to intervention and a critical target for therapeutic breakthroughs. Although microbiome-targeted approaches, such as FMT and probiotics, provide promising routes for addressing the underlying etiology, they remain largely exploratory, and current evidence regarding clinical efficacy and safety is still inadequate. Accordingly, future research should prioritize two key areas: first, the development of novel assessment tools that integrate dynamic measures of metabolism, inflammation, and body composition to enable early and precise identification of nutritional risk; second, the design and conduct of large-scale prospective intervention studies with clinically meaningful hard endpoints - such as muscle mass and survival - to clarify the role of microbiome modulation within comprehensive treatment strategies supported by robust evidence, ultimately improving outcomes for this expanding patient population.
DECLARATIONS
Authors’ contributions
Made contributions to the conception of the study: Huang BZ
Collected and analyzed the literature: Huang BZ
Contributed to manuscript discussion: Tian CB
Revised the manuscript for final submission: Huang BZ
Writing - review & editing: Sui YT
Availability of data and materials
Not applicable.
AI and AI-assisted tools statement
During the preparation of this manuscript, the AI tool Tencent Yuanbao (version 2.72.0, released 2026-06-12) was used solely for language editing. The tool did not influence the study design, data collection, analysis, interpretation, or the scientific content of the work. All authors take full responsibility for the accuracy, integrity, and final content of the manuscript.
Financial support and sponsorship
This work was supported by the Shenzhen Basic Research Special Project (Grant No. JCYJ20240813145222029).
Conflicts of interest
All authors declared that there are no conflicts of interest.
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Copyright
© The Author(s) 2026.
REFERENCES
1. Li P, Ding Z, Feng Y, et al. Global, regional, and national burden of hepatocellular carcinoma and contribution of nine modifiable risk factors across 185 countries/territories in 2022. Sci Bull. 2026;71:838-49.
2. Chong B, Kong G, Shankar K, et al. The global syndemic of metabolic diseases in the young adult population: a consortium of trends and projections from the Global Burden of Disease 2000-2019. Metabolism. 2023;141:155402.
3. Riazi K, Azhari H, Charette JH, et al. The prevalence and incidence of NAFLD worldwide: a systematic review and meta-analysis. Lancet Gastroenterol Hepatol. 2022;7:851-61.
4. Na BG, Han SS, Cho YA, et al. Nutritional status of patients with cancer: a prospective cohort study of 1,588 hospitalized patients. Nutr Cancer. 2018;70:1228-36.
5. Kim Y, Kim SR, Kim K, Yu SJ. Demographic, clinical and psychological predictors of malnutrition among people with liver cancer. Eur J Oncol Nurs. 2024;68:102497.
6. Rios TC, de Oliveira LPM, da Costa MLV, et al. A poorer nutritional status impacts quality of life in a sample population of elderly cancer patients. Health Qual Life Outcomes. 2021;19:90.
7. da Silva GD, Batista AVA, Costa MCRA, Dos Santos ACO. The ability of GLIM and MNA-FF to diagnose malnutrition and predict sarcopenia and frailty in hospitalized adults over 60 years of age. Front Nutr. 2024;11:1456091.
8. Xu Y, Hao X, Ren Y, et al. Research progress of abnormal lactate metabolism and lactate modification in immunotherapy of hepatocellular carcinoma. Front Oncol. 2022;12:1063423.
9. Lequoy M, Gigante E, Couty JP, Desbois-Mouthon C. Hepatocellular carcinoma in the context of non-alcoholic steatohepatitis (NASH): recent advances in the pathogenic mechanisms. Horm Mol Biol Clin Investig. 2020;41:1-44.
10. Renehan AG, Zwahlen M, Egger M. Adiposity and cancer risk: new mechanistic insights from epidemiology. Nat Rev Cancer. 2015;15:484-98.
11. Ruan X, Nakyeyune R, Shao Y, et al. Nutritional screening tools for adult cancer patients: a hierarchical Bayesian latent-class meta-analysis. Clin Nutr. 2021;40:1733-43.
12. Xie H, Yuan K, Ruan G, et al. Improving the assessment of malnutrition in cancer: Using systemic inflammation markers as a supplement to the inflammation items of the GLIM criteria. Clin Nutr. 2023;42:2036-44.
13. Gort-van Dijk D, Weerink LBM, Milovanovic M, et al. Bioelectrical impedance analysis and mid-upper arm muscle circumference can be used to detect low muscle mass in clinical practice. Nutrients. 2021;13:2350.
14. Kotoh Y, Saeki I, Yamasaki T, et al. Albumin-bilirubin score as a useful predictor of energy malnutrition in patients with hepatocellular carcinoma. Clin Nutr. 2021;40:3585-91.
15. Bischoff SC, Bernal W, Dasarathy S, et al. ESPEN practical guideline: clinical nutrition in liver disease. Clin Nutr. 2020;39:3533-62.
16. Trujillo EB, Kadakia KC, Thomson C, et al. Malnutrition risk screening in adult oncology outpatients: an ASPEN systematic review and clinical recommendations. JPEN J Parenter Enteral Nutr. 2024;48:874-94.
17. Ge Y, Shi H, shen X. Aiww: a new nutrition screening tool for the oncologic population. Clinical Nutrition ESPEN. 2023;54:541.
18. Wiegert EVM, Padilha PC, Peres WAF. Performance of patient-generated subjective global assessment (PG-SGA) in patients with advanced cancer in palliative care. Nutr Clin Pract. 2017;32:675-81.
19. Zhang KP, Tang M, Fu ZM, et al. Global Leadership Initiative on Malnutrition criteria as a nutrition assessment tool for patients with cancer. Nutrition. 2021;91-92:111379.
20. Rosnes KS, Henriksen C, Høidalen A, Paur I. Agreement between the GLIM criteria and PG-SGA in a mixed patient population at a nutrition outpatient clinic. Clin Nutr. 2021;40:5030-7.
21. Sobrini P, Sánchez-Castellano C, Cruz-Jentoft AJ. MNA-SF as a screening tool for malnutrition diagnosed with the glim criteria in older persons with cancer. Eur Geriatr Med. 2021;12:653-6.
22. McFarlane M, Hammond C, Roper T, et al. Comparing assessment tools for detecting undernutrition in patients with liver cirrhosis. Clin Nutr ESPEN. 2018;23:156-61.
23. Borhofen SM, Gerner C, Lehmann J, et al. The royal free hospital-nutritional prioritizing tool is an independent predictor of deterioration of liver function and survival in cirrhosis. Dig Dis Sci. 2016;61:1735-43.
24. Georgiou A, Papatheodoridis GV, Alexopoulou A, et al. Evaluation of the effectiveness of eight screening tools in detecting risk of malnutrition in cirrhotic patients: the KIRRHOS study. Br J Nutr. 2019;122:1368-76.
25. Xu J, Li J, Chen H, et al. Prevalence and prognostic significance of malnutrition in patients with non-alcoholic fatty liver disease (NAFLD). Int J Vitam Nutr Res. 2024;95:26099.
26. Ruiz-Margáin A, Román-Calleja BM, Moreno-Guillén P, et al. Nutritional therapy for hepatocellular carcinoma. World J Gastrointest Oncol. 2021;13:1440-52.
27. Topan MM, Sporea I, Dănilă M, et al. Comparison of different nutritional assessment tools in detecting malnutrition and sarcopenia among cirrhotic patients. Diagnostics. 2022;12:893.
28. Sguanci M, Palomares SM, Cangelosi G, et al. Artificial intelligence in the management of malnutrition in cancer patients: a systematic review. Adv Nutr. 2025;16:100438.
29. Zhang Z, Wan Z, Zhu Y, Zhang L, Zhang L, Wan H. Prevalence of malnutrition comparing NRS2002, MUST, and PG-SGA with the GLIM criteria in adults with cancer: a multi-center study. Nutrition. 2021;83:111072.
30. Zhang P, Wang Q, Zhu M, Li P, Wang Y. Differences in nutritional risk assessment between NRS2002, RFH-NPT and LDUST in cirrhotic patients. Sci Rep. 2023;13:3306.
31. Rajab N, Abdul Hamid SB, Mohd Said AH, Md Isa KA. Validation of nutrition screening tool: royal free hospital nutritional prioritizing tool (RFH-NPT) for chronic liver disease patients. MJMHS. 2023;19:130-7.
32. Wang R, Huang L, Xu M, Yu X, Wang H. Comparison of different nutritional screening tools in nutritional screening of patients with cirrhosis: a cross-sectional observational study. Heliyon. 2024;10:e30339.
33. Liu CH, Zeng QM, Kim W, et al. Sarcopenia and MASLD: novel insights and the future. Nat Rev Endocrinol. 2026;22:139-52.
34. Kim G, Lee SE, Lee YB, et al. Relationship between relative skeletal muscle mass and nonalcoholic fatty liver disease: a 7-year longitudinal study. Hepatology. 2018;68:1755-68.
35. Petermann-Rocha F, Gray SR, Forrest E, et al. Associations of muscle mass and grip strength with severe NAFLD: a prospective study of 333,295 UK Biobank participants. J Hepatol. 2022;76:1021-9.
36. Ponziani FR, Bhoori S, Castelli C, et al. Hepatocellular carcinoma is associated with gut microbiota profile and inflammation in nonalcoholic fatty liver disease. Hepatology. 2019;69:107-20.
37. Selvam S, Vairappan B. Tight junction proteins: gatekeepers turned facilitators in the pathogenesis of gastric adenocarcinoma. World J Gastrointest Oncol. 2026;18:114040.
38. Wang X, Li MM, Niu Y, et al. Serum zonulin in HBV-associated chronic hepatitis, liver cirrhosis, and hepatocellular carcinoma. Dis Markers. 2019;2019:5945721.
39. Kolodziejczyk AA, Zheng D, Shibolet O, Elinav E. The role of the microbiome in NAFLD and NASH. EMBO Mol Med. 2019;11:e9302.
40. Dong YQ, Lu CW, Zhang L, Yang J, Hameed W, Chen W. Toll-like receptor 4 signaling promotes invasion of hepatocellular carcinoma cells through MKK4/JNK pathway. Mol Immunol. 2015;68:671-83.
41. Che Z, Xue W, Zhao X, Hu C, Tian Y. Regulatory role and biomarker potential of gut microbiota metabolites in the progression of metabolic dysfunction-associated steatotic liver disease to hepatocellular carcinoma. Clin Transl Gastroenterol. 2025;16:e00914.
42. Pierantonelli I, Svegliati-Baroni G. Nonalcoholic fatty liver disease: basic pathogenetic mechanisms in the progression from NAFLD to NASH. Transplantation. 2019;103:e1-13.
43. Chang KV, Wu WT, Chen YH, et al. Enhanced serum levels of tumor necrosis factor-α, interleukin-1β, and -6 in sarcopenia: alleviation through exercise and nutrition intervention. Aging. 2023;15:13471-85.
44. Choi K, Jang HY, Ahn JM, et al. The association of the serum levels of myostatin, follistatin, and interleukin-6 with sarcopenia, and their impacts on survival in patients with hepatocellular carcinoma. Clin Mol Hepatol. 2020;26:492-505.
45. Ding J, Yang G, Sun W, et al. Association of interleukin-6 with sarcopenia and its components in older adults: a systematic review and meta-analysis of cross-sectional studies. Ann Med. 2024;56:2384664.
46. Johnson DE, O’Keefe RA, Grandis JR. Targeting the IL-6/JAK/STAT3 signalling axis in cancer. Nat Rev Clin Oncol. 2018;15:234-48.
47. Jaśkiewicz A, Domoradzki T, Pająk B. Targeting the JAK2/STAT3 pathway-can we compare it to the two faces of the god janus? Int J Mol Sci 2020;21:8261.
48. Sciorati C, Gamberale R, Monno A, et al. Pharmacological blockade of TNFα prevents sarcopenia and prolongs survival in aging mice. Aging. 2020;12:23497-508.
49. Yi X, Tao J, Qian Y, et al. Morroniside ameliorates inflammatory skeletal muscle atrophy via inhibiting canonical and non-canonical NF-κB and regulating protein synthesis/degradation. Front Pharmacol. 2022;13:1056460.
50. Li J, Yi X, Yao Z, Chakkalakal JV, Xing L, Boyce BF. TNF receptor-associated factor 6 mediates TNFα-induced skeletal muscle atrophy in mice during aging. J Bone Miner Res. 2020;35:1535-48.
51. Lahiri S, Kim H, Garcia-Perez I, et al. The gut microbiota influences skeletal muscle mass and function in mice. Sci Transl Med. 2019;11:eaan5662.
52. Li L, Lian P, Dong W, et al. Restoring muribaculum intestinale-derived butyrate mitigates skeletal muscle loss in cancer cachexia. J Cachexia Sarcopenia Muscle. 2025;16:e70140.
53. Yoshida H, Ishii M, Akagawa M. Propionate suppresses hepatic gluconeogenesis via GPR43/AMPK signaling pathway. Arch Biochem Biophys. 2019;672:108057.
54. Asaoka D, Toda K, Yoshimoto S, et al. Sex-specific associations of gut microbiota composition with sarcopenia defined by the Asian working group for sarcopenia 2019 consensus in older outpatients: prospective cross-sectional study in Japan. Nutrients. 2025;17:1746.
55. Liu C, Cheung WH, Li J, et al. Understanding the gut microbiota and sarcopenia: a systematic review. J Cachexia Sarcopenia Muscle. 2021;12:1393-407.
57. Ippolito L, Morandi A, Giannoni E, Chiarugi P. Lactate: A metabolic driver in the tumour landscape. Trends Biochem Sci. 2019;44:153-66.
58. Lin J, Rao D, Zhang M, Gao Q. Metabolic reprogramming in the tumor microenvironment of liver cancer. J Hematol Oncol. 2024;17:6.
59. Órdenes P, Villar PS, Tarifeño-Saldivia E, et al. Lactate activates hypothalamic POMC neurons by intercellular signaling. Sci Rep. 2021;11:21644.
60. Liu X, Li S, Cui Q, et al. Activation of GPR81 by lactate drives tumour-induced cachexia. Nat Metab. 2024;6:708-23.
61. Davis MP, Dreicer R, Walsh D, Lagman R, LeGrand SB. Appetite and cancer-associated anorexia: a review. J Clin Oncol. 2004;22:1510-7.
62. Han J, Liang X, Guo Y, Wu X, Li Z, Hong T. Agouti-related protein as the glucose signaling sensor in the central melanocortin circuits in regulating fish food intake. Front Endocrinol. 2022;13:1010472.
63. Yeom E, Yu K. Understanding the molecular basis of anorexia and tissue wasting in cancer cachexia. Exp Mol Med. 2022;54:426-32.
64. Laird BJ, McMillan D, Skipworth RJE, et al. The emerging role of interleukin 1β (IL-1β) in cancer cachexia. Inflammation. 2021;44:1223-8.
65. Braun TP, Zhu X, Szumowski M, et al. Central nervous system inflammation induces muscle atrophy via activation of the hypothalamic-pituitary-adrenal axis. J Exp Med. 2011;208:2449-63.
66. Campos CA, Bowen AJ, Han S, Wisse BE, Palmiter RD, Schwartz MW. Cancer-induced anorexia and malaise are mediated by CGRP neurons in the parabrachial nucleus. Nat Neurosci. 2017;20:934-42.
67. Roman CW, Derkach VA, Palmiter RD. Genetically and functionally defined NTS to PBN brain circuits mediating anorexia. Nat Commun. 2016;7:11905.
68. Olson B, Zhu X, Norgard MA, et al. Lipocalin 2 mediates appetite suppression during pancreatic cancer cachexia. Nat Commun. 2021;12:2057.
69. Li X, Holtrop T, Jansen FAC, et al. Lipopolysaccharide-induced hypothalamic inflammation in cancer cachexia-anorexia is amplified by tumour-derived prostaglandin E2. J Cachexia Sarcopenia Muscle. 2022;13:3014-27.
70. Cao HM, Ye XP, Ma JH, et al. Mimecan, a hormone abundantly expressed in adipose tissue, reduced food intake independently of leptin signaling. EBioMedicine. 2015;2:1718-24.
71. Borner T, Liberini CG, Lutz TA, Riediger T. Brainstem GLP-1 signalling contributes to cancer anorexia-cachexia syndrome in the rat. Neuropharmacology. 2018;131:282-90.
72. Liu J, Jing C, Guo Y, et al. The central signaling pathways related to metabolism-regulating hormones of the gut-brain axis: a review. J Transl Med. 2025;23:648.
73. Tolhurst G, Heffron H, Lam YS, et al. Short-chain fatty acids stimulate glucagon-like peptide-1 secretion via the G-protein-coupled receptor FFAR2. Diabetes. 2012;61:364-71.
74. Nøhr MK, Egerod KL, Christiansen SH, et al. Expression of the short chain fatty acid receptor GPR41/FFAR3 in autonomic and somatic sensory ganglia. Neuroscience. 2015;290:126-37.
75. Martin CR, Osadchiy V, Kalani A, Mayer EA. The brain-gut-microbiome axis. Cell Mol Gastroenterol Hepatol. 2018;6:133-48.
76. Sanchez JG, Enriquez JR, Wells JM. Enteroendocrine cell differentiation and function in the intestine. Curr Opin Endocrinol Diabetes Obes. 2022;29:169-76.
77. Duran M, Willis JR, Dalvi N, Fokakis Z, Virkus SA, Hardaway JA. Integration of glucagon-like peptide 1 receptor actions through the central amygdala. Endocrinology. 2025;166:bqaf019.
78. Al-Amouri FM, Badrasawi M. Taste alteration and its relationship with nutritional status among cancer patients receiving chemotherapy, cross-sectional study. PLoS One. 2024;19:e0302990.
79. Fons A, Oreskovich D, Prioult G, von der Weid B, Lerea-Antes J. Taste alterations in patients with cancer: an overview and recommendations for future research. Support Care Cancer. 2025;33:784.
80. Juthani R, Malalur P, Manne A, Mittra A. The combined use of lenvatinib and locoregional therapies for the management of hepatocellular carcinoma. Cancers. 2025;17:1572.
81. Hustad KS, Koteng LH, Urrizola A, et al. ; MyPath consortium. Practical cancer nutrition, from guidelines to clinical practice: a digital solution to patient-centred care. ESMO Open. 2025;10:104529.
82. Vaughn BP, Rank KM, Khoruts A. Fecal microbiota transplantation: current status in treatment of GI and liver disease. Clin Gastroenterol Hepatol. 2019;17:353-61.
83. Ademe M. Benefits of fecal microbiota transplantation: a comprehensive review. J Infect Dev Ctries. 2020;14:1074-80.
84. Gu C, Sha G, Zeng B, Cao H, Cao Y, Tang D. Therapeutic potential of fecal microbiota transplantation in colorectal cancer based on gut microbiota regulation: from pathogenesis to efficacy. Ther Adv Gastroenterol. 2025;18:17562848251327167.
85. Lim MY, Hong S, Nam YD. Understanding the role of the gut microbiome in solid tumor responses to immune checkpoint inhibitors for personalized therapeutic strategies: a review. Front Immunol. 2024;15:1512683.
86. Karimi M, Shirsalimi N, Hashempour Z, et al. Safety and efficacy of fecal microbiota transplantation (FMT) as a modern adjuvant therapy in various diseases and disorders: a comprehensive literature review. Front Immunol. 2024;15:1439176.
87. Yong D. Screening for potential infections in fecal microbiota transplantation guidelines and an experience of microbiota bank operation in Korea: a narrative review. Ann Clin Microbiol. 2024;27:11-7.
88. Fuhri Snethlage CM, Nieuwdorp M, Hanssen NMJ. Faecal microbiota transplantation in endocrine diseases and obesity. Best Pract Res Clin Endocrinol Metab. 2021;35:101483.
89. Elghannam MT, Hassanien MH, Ameen YA, et al. Gut microbiome and gastric cancer: microbial interactions and therapeutic potential. Gut Pathog. 2025;17:56.
90. Kao D, Roach B, Silva M, et al. Effect of oral capsule- vs colonoscopy-delivered fecal microbiota transplantation on recurrent clostridium difficile infection: a randomized clinical trial. JAMA. 2017;318:1985-93.
91. Baxter M, Colville A. Adverse events in faecal microbiota transplant: a review of the literature. J Hosp Infect. 2016;92:117-27.
92. Yadegar A, Bar-Yoseph H, Monaghan TM, et al. Fecal microbiota transplantation: current challenges and future landscapes. Clin Microbiol Rev. 2024;37:e0006022.
93. Xiao Y, Angulo MT, Lao S, Weiss ST, Liu YY. An ecological framework to understand the efficacy of fecal microbiota transplantation. Nat Commun. 2020;11:3329.
94. Swanson KS, Gibson GR, Hutkins R, et al. The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of synbiotics. Nat Rev Gastroenterol Hepatol. 2020;17:687-701.
95. Han SH, Suk KT, Kim DJ, et al. Effects of probiotics (cultured Lactobacillus subtilis/Streptococcus faecium) in the treatment of alcoholic hepatitis: randomized-controlled multicenter study. Eur J Gastroenterol Hepatol. 2015;27:1300-6.
96. Zhao Z, Chen L, Zhao Y, et al. Lactobacillus plantarum NA136 ameliorates nonalcoholic fatty liver disease by modulating gut microbiota, improving intestinal barrier integrity, and attenuating inflammation. Appl Microbiol Biotechnol. 2020;104:5273-82.
97. Manzhalii E, Virchenko O, Falalyeyeva T, Beregova T, Stremmel W. Treatment efficacy of a probiotic preparation for non-alcoholic steatohepatitis: a pilot trial. J Dig Dis. 2017;18:698-703.
98. Sepideh A, Karim P, Hossein A, et al. Effects of multistrain probiotic supplementation on glycemic and inflammatory indices in patients with nonalcoholic fatty liver disease: a double-blind randomized clinical trial. J Am Coll Nutr. 2016;35:500-5.
99. Min S, Than N, Shin YC, et al. Live probiotic bacteria administered in a pathomimetic Leaky Gut Chip ameliorate impaired epithelial barrier and mucosal inflammation. Sci Rep. 2022;12:22641.
100. Ye Z, Tan Q, Woltemate S, et al. Escherichia coli nissle improves short-chain fatty acid absorption and barrier function in a mouse model for chronic inflammatory diarrhea. Inflamm Bowel Dis. 2025;31:1109-20.
101. Roobab U, Batool Z, Manzoor MF, Shabbir MA, Khan MR, Aadil RM. Sources, formulations, advanced delivery and health benefits of probiotics. Current Opinion in Food Science. 2020;32:17-28.
102. Silva DR, Sardi JDCO, Pitangui NDS, Roque SM, Silva ACBD, Rosalen PL. Probiotics as an alternative antimicrobial therapy: current reality and future directions. J Funct Foods. 2020;73:104080.
103. Akbar Z, Zahoor T, Huma N, Jamil A, Ayesha H, Kumar Irudayaraj JM. Electrospun probiotics: an alternative for encapsulation. J Biol Regul Homeost Agents. 2018;32:1551-6.
104. Huang Y, Zhang L, Hu J, Liu H. Improved loading capacity and viability of probiotics encapsulated in alginate hydrogel beads by in situ cultivation method. Foods. 2023;12:2256.
105. Liu Q, Lin C, Yang X, et al. Improved viability of probiotics via microencapsulation in Whey-Protein-Isolate-Octenyl-Succinic-Anhydride-Starch-Complex coacervates. Molecules. 2023;28:5732.
106. Albadran HA, Monteagudo-Mera A, Khutoryanskiy VV, Charalampopoulos D. Development of chitosan-coated agar-gelatin particles for probiotic delivery and targeted release in the gastrointestinal tract. Appl Microbiol Biotechnol. 2020;104:5749-57.
107. Fu Y, Liu L, Zhang J, et al. Reinforcing alginate matrixes by tea polysaccharide conjugates or their stabilized nanoemulsion for probiotics encapsulation: characterization, survival after gastrointestinal digestion and ambient storage. Int J Biol Macromol. 2023;253:126828.
108. Hao R, Chen Z, Wu Y, et al. Improving the survival of Lactobacillus plantarum FZU3013 by phase separated caseinate/alginate gel beads. Int J Biol Macromol. 2024;260:129447.
Cite This Article
How to Cite
Download Citation
Export Citation File:
Type of Import
Tips on Downloading Citation
Citation Manager File Format
Type of Import
Direct Import: When the Direct Import option is selected (the default state), a dialogue box will give you the option to Save or Open the downloaded citation data. Choosing Open will either launch your citation manager or give you a choice of applications with which to use the metadata. The Save option saves the file locally for later use.
Indirect Import: When the Indirect Import option is selected, the metadata is displayed and may be copied and pasted as needed.
About This Article
Special Topic
Copyright

Data & Comments
Data
0












Comments
Comments must be written in English. Spam, offensive content, impersonation, and private information will not be permitted. If any comment is reported and identified as inappropriate content by OAE staff, the comment will be removed without notice. If you have any queries or need any help, please contact us at support@oaepublish.com.