


Prussian blue nanozyme for ischemia-reperfusion injury treatment: progress, opportunities and challenges
 0
0 Abstract
Ischemia-reperfusion injury (IRI) is a pathophysiological process occurring after blood supply restoration to tissues or organs, causing unexpected cellular damage. It involves multiple complex mechanisms, but inflammation and oxidative stress are the main causes. IRI can damage vital organs such as the heart, brain, liver, and kidneys, affecting patient prognosis and quality of life. Therefore, the prevention and treatment of IRI have become a key area in clinical and basic research. Prussian blue, a metal-organic framework material, is approved by the U.S. Food and Drug Administration for treating heavy metal poisoning. In 2016, research indicated that Prussian blue nanoparticles had multi-enzyme activities like catalase, peroxidase, and superoxide dismutase, enabling them to scavenge reactive oxygen species and combat inflammation, establishing them as a highly promising nanozyme. Leveraging their unique antioxidative and anti-inflammatory properties, Prussian blue nanozyme (PBNZ) can directly target key pathological pathways of IRI. Furthermore, PBNZ can serve as efficient drug delivery systems, and through functional modifications, they enable the targeted delivery of therapeutic agents, thereby synergistically enhancing treatment efficacy. Currently, PBNZ have demonstrated significant therapeutic potential in IRI models involving various organs and tissues such as the heart, brain, liver, kidneys, and skin flaps. This review aims to outline IRI's key pathological mechanisms, analyze how PBNZ combats oxidative stress and inflammation, and summarize its recent application advancements in treating IRI in different organs. The goal is to offer theoretical reference and research insights for the future development of novel nanozyme-based therapeutic strategies against IRI.
Keywords
INTRODUCTION
Ischemia-reperfusion injury (IRI) is a complex pathophysiological phenomenon, which is defined as follows: after a period of ischemia in tissue or an organ, even if blood flow is restored, the reperfusion itself can lead to further exacerbation of tissue damage[1]. IRI primarily consists of two key stages: the "ischemic" phase and the "reperfusion" phase[2]. During the "ischemic" phase, tissues experience disturbances in energy metabolism due to the lack of oxygen and nutrients, which in turn triggers a series of cellular dysfunctions and may ultimately lead to cell death[3]. While in the "Reperfusion" phase, although the ischemic tissue regains blood flow and oxygen supply, the tissue damage may show a progressive intensification[4]. Studies have shown that the sudden influx of oxygen during reperfusion triggers oxidative stress and leads to excessive production of reactive oxygen species (ROS)[5]. Additionally, reperfusion activates the immune system and initiates a robust inflammatory response[5]. Ultimately, this cascade of events worsens cellular dysfunction and imposes additional stress on already compromised ischemic tissue, creating a vicious cycle. IRI commonly occurs in clinical settings such as myocardial infarction, stroke reperfusion therapy, and organ transplantation[6]. It is closely associated with damage to vital organs, including the heart, liver, lungs, kidneys, and brain[3,7-10], significantly affecting treatment outcomes and prognosis. Therefore, IRI represents a critical clinical challenge that demands urgent attention. A deeper understanding of its pathophysiological mechanisms and the development of novel therapeutic strategies are of great importance for improving the diagnosis and treatment of related diseases.
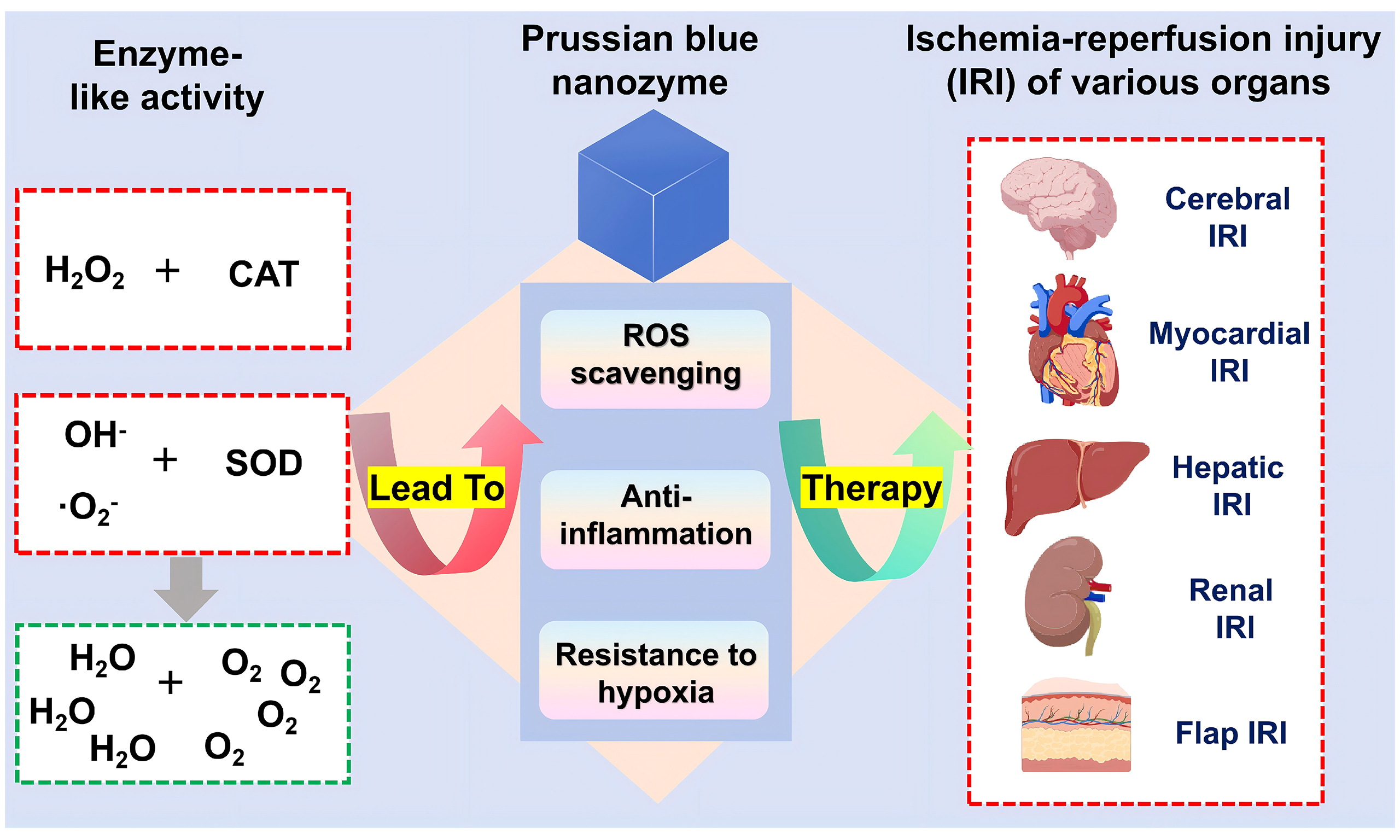
Prussian blue (PB) is a metal-organic framework (MOF) with a unique structure composed of alternating iron ions (Fe3+ and Fe2+) coordinated with cyanide, which is a time-honored deep blue pigment and also a material with good safety for human use[11]. In 2003, the U.S. Food and Drug Administration officially approved soluble PB for the treatment of heavy metal thallium and cesium poisoning[12]. With the continuous innovation of nanoscience and nanotechnology, PB has been successfully prepared into Prussian blue nanoparticles (PBNPs) with nanoscale dimensions, and PBNPs exhibit unique properties and excellent biocompatibility, and are widely used in the biomedical field[12]. In 2016, Zhang and others first discovered that PBNPs have enzyme-like activities similar to catalase (CAT), peroxidase (POD), and superoxide dismutase (SOD), which can effectively scavenge ROS and combat inflammation[13]. Since then, PBNPs have entered the field of nanozymes, and their unique multi-enzyme activities enable them to play a wide range of roles in the biomedical field, providing innovative strategies for the treatment of tumors, cardiovascular diseases, inflammatory bowel diseases, wound healing, etc.[14].
Inflammatory infiltration and oxidative stress are the most prominent pathological features of IRI and serve as critical therapeutic targets in the development of IRI treatment strategies. Prussian Blue Nanozyme (PBNZ), owing to its unique multi-enzyme-like properties, has demonstrated remarkable capabilities in scavenging ROS and exerting anti-inflammatory effects[13], making it a highly promising therapeutic approach for IRI. Additionally, PBNZ can be engineered as an efficient drug delivery system. Through rational structural optimization and functional modifications, it enables targeted delivery of therapeutic agents with low bioavailability or poor solubility[15], thereby further enhancing the therapeutic efficacy of IRI. In recent years, numerous studies have reported the extensive application of PBNZ in IRI treatment, covering multiple vital organs such as the brain[16], heart[17], liver[18], kidneys[19], and flaps[20], fully demonstrating its adaptability and therapeutic potential across diverse pathological environments.
Against this backdrop, this review aims to provide a comprehensive and systematic summary of the latest research progress on PBNZ in the treatment of IRI. First, we will delve into the core mechanisms underlying the development of IRI, with a particular focus on the specific roles and interrelationships of oxidative stress and inflammatory infiltration—two key pathological processes. Subsequently, we will detail the antioxidative stress and anti-inflammatory mechanisms of PBNZ, highlighting its unique advantages in mitigating IRI-related pathological changes. Finally, this article will categorize and review the specific therapeutic effects of PBNZ in IRI treatment across different tissues and organs (e.g., heart, brain, liver, kidneys, and flaps), supported by detailed analyses of existing experimental data and research findings. Through this systematic review and synthesis, we hope to offer valuable insights and inspiration for future research on PBNZ and other nanomaterials in the field of IRI treatment, while also laying a theoretical foundation to further advance developments in this area.
INFLAMMATION AND OXIDATIVE STRESS OF IRI
IRI is a common complication in diverse clinical settings, including thrombolytic therapy for myocardial or cerebral infarction, organ transplantation, and the repair of traumatic limb injuries[21]. It can result in severe clinical consequences such as acute heart failure, neurological dysfunction, systemic inflammatory response syndrome (SIRS), and multiple organ dysfunction syndrome (MODS)[5,22]. Based on the affected tissues or organs, IRI can be classified into several types, such as myocardial IRI, cerebral IRI, hepatic IRI, renal IRI, intestinal IRI, and flap IRI[6]. Each type of IRI has its unique pathophysiological mechanisms and clinical manifestations, yet all involve core pathological processes including oxidative stress and inflammatory infiltration[23].
Inflammation
The inflammatory response in IRI serves as a central driver of tissue damage, marked by its cascade-amplifying and destructive nature. This process begins during the ischemic phase, where cells under ischemia experience depletion of cellular energy (ATP), acidosis, and ion imbalance due to the lack of oxygen and nutrients. These internal environmental disturbances trigger intense intracellular stress signals and promote the release of Damage-Associated Molecular Patterns (DAMPs), thereby initiating the adaptive immune response[4,24]. Hypoxia also stabilizes and activates Hypoxia-Inducible Factor-1α (HIF-1α), which can directly upregulate the expression of various pro-inflammatory factors and chemokines such as inducible Nitric Oxide Synthase (iNOS), Interleukin-1β (IL-1β), Tumor Necrosis Factor-α (TNF-α), and Monocyte Chemoattractant Protein-1 (MCP-1)[25], creating a chemical gradient for subsequent leukocyte recruitment and inflammation. Therefore, the inflammatory response during ischemia is a relatively "quiet" yet crucial warning and preparatory stage. Through metabolic reprogramming and epigenetic changes, it paves the way for the devastating "inflammatory storm" that occurs during the reperfusion phase[26].
Upon entering the reperfusion phase, the explosive generation of ROS and calcium overload further activates endothelial cells and the innate immune system, rapidly triggering a series of intense, explosive, and destructive inflammatory reactions[27]. These reactions involve not only the participation of various immune cells but also the release of a large number of inflammatory mediators and the activation of signaling pathways. Specifically, vascular endothelial cells swiftly surface-express P-selectin and E-selectin, which mediate the adhesion, rolling, and migration of inflammatory cells along the endothelium, facilitating their recruitment to the site of injury and amplifying the inflammatory process[28]. Subsequently, under IRI stimulation, vascular endothelial cells express adhesion molecules such as Intercellular Adhesion Molecule-1 (ICAM-1) and Vascular Cell Adhesion Molecule-1 (VCAM-1)[29]. These adhesion molecules bind to corresponding receptors on the surface of neutrophils, promoting neutrophil adhesion to the endothelium and their subsequent transmigration through the vessel wall into the tissue[30]. The recruited neutrophils can release various active substances, including proteases, ROS, and pro-inflammatory cytokines (such as TNF-α, IL-1β, and IL-6). These substances damage microvascular endothelial cells, increase microvascular permeability, cause plasma component exudation, and lead to tissue edema[31,32]. Concurrently, the NLRP3 inflammasome is activated, catalyzing the maturation of inflammatory cytokines IL-1β and IL-18 and inducing pyro ptosis, thereby further amplifying inflammation[33]. In addition, IRI can activate complement components via the classical, alternative, and lectin pathways[34]. Upon activation, the complement system elicits diverse biological effects; for instance, both C3a and C5a exert chemotactic activity, recruiting inflammatory cells such as neutrophils and monocytes[34,35]. These inflammatory mediators exacerbate the local tissue inflammatory response, forming a positive feedback loop. This leads to loss of cell membrane integrity, mitochondrial dysfunction, increased apoptosis, and ultimately results in extensive cellular damage and organ dysfunction. The excessive inflammatory response not only directly kills parenchymal cells but also exacerbates tissue injury by causing microvascular dysfunction and the "no-reflow" phenomenon.
Oxidative stress
Oxidative stress is a critical mechanism underlying IRI, with its induction in IRI attributed to several key processes. During the ischemic stage, insufficient blood supply disrupts oxygen-dependent metabolism, impairing mitochondrial oxidative phosphorylation and drastically reducing ATP synthesis[2]. ATP is sequentially degraded to ADP, AMP, adenosine, and hypoxanthine, leading to the accumulation of hypoxanthine[36]. Concurrently, intracellular calcium overload activates calcium-dependent proteases, which convert xanthine dehydrogenase (XD) to xanthine oxidase (XO) in vascular endothelial cells[36]. By this stage, both XO and its substrate hypoxanthine are accumulated, priming the system for an oxidative burst upon reperfusion.
Upon reperfusion and reoxygenation, molecular oxygen (O2) is delivered to ischemic tissues, serving as the critical substrate for XO. XO catalyzes the oxidation of hypoxanthine to xanthine and further to uric acid, transferring electrons to O2 during these reactions to generate large quantities of superoxide anion (•O2-) and hydrogen peroxide (H2O2),[37] which are the primary sources of ROS in early reperfusion. Ischemic damage also impairs the mitochondrial electron transport chain (ETC) complexes. During reperfusion, influx of O2 leads to "single-electron leakage" from dysfunctional ETC complexes, causing incomplete oxygen reduction and sustained •O2-[38], representing a persistent source of ROS. Additionally, early ROS and post-reperfusion chemokines (e.g., leukotriene B4) synergistically activate and recruit neutrophils to the injury site. Activated neutrophils express NADPH oxidase (NOX) on their membranes, which undergoes a "respiratory burst" to consume oxygen and generate •O2-[39]. ROS further propagate injury by attacking membrane polyunsaturated fatty acids, triggering lipid peroxidation cascades. This generates additional free radicals and oxidative end products (e.g., malondialdehyde, 4-hydroxynonenal), amplifying oxidative stress[40,41].
Notably, ischemic energy depletion compromises the activity of intrinsic antioxidant enzymes (e.g., SOD, CAT and glutathione peroxidase) and non-enzymatic antioxidants (e.g., glutathione, vitamins C/E), which significantly attenuates ROS-scavenging capacity[42]. In summary, while reperfusion restores oxygen supply, it fails to restore antioxidant function at a rate commensurate with the explosive generation of ROS. This imbalance culminates in oxidative stress dysregulation, which damages cellular components—including membranes, proteins, and DNA—ultimately leading to cell death.
During the onset and progression of IRI, oxidative stress and inflammatory infiltration operate in a synergistic manner. Specifically, the excessive ROS generated under oxidative stress not only directly damage critical cellular structures—including membranes, proteins, and nucleic acids—but also activate complex signaling pathways such as nuclear factor kappa B (NF-κB), thereby inducing the release of various pro-inflammatory cytokines and further exacerbating local tissue inflammation.[22] This ROS-triggered release of inflammatory factors establishes a positive feedback loop wherein oxidative stress and inflammation mutually reinforce each other, continuously amplifying tissue injury. Concurrently, extensive inflammatory cell infiltration leads to the secretion of various enzymes, with myeloperoxidase (MPO) serving as a prominent example[43]. These enzymes catalyze the generation of additional ROS, further elevating free radical concentrations and intensifying oxidative damage. The newly generated ROS, in turn, stimulate further inflammatory factor release, creating a self-perpetuating cycle of pathological amplification. Through this dual-hit mechanism, cellular damage in ischemic tissues—which may initially be reversible upon reperfusion—progressively transitions toward irreversible necrosis. This progression ultimately results in significant functional decline or even complete loss of organ function, posing a serious threat to patient health. Consequently, both the inflammatory response and oxidative stress represent critical therapeutic targets in IRI, and interventions aimed at mitigating these processes hold promise for effectively reversing IRI-induced cell damage.
ANTIOXIDANT STRESS AND ANTI-INFLAMMATION EFFECTS OF PBNZ
PBNZ, as a class of emerging nanomaterials, have garnered significant attention in the biomedical field in recent years. They possess both the unique chemical properties of Prussian blue and the highly efficient catalytic activity of nanozymes. Their catalytic activity can be modulated by adjusting parameters such as size, crystallinity, morphology, and surface modifications, enabling them to meet diverse application requirements[14]. PBNZ can mimic the catalytic functions of natural enzymes, efficiently catalyzing specific chemical reactions under mild conditions. Leveraging their unique enzyme-like activities, PBNZ can efficiently scavenge excess ROS within the body, thereby mitigating the damage caused by oxidative stress to cells and tissues. Concurrently, PBNZ can exert significant anti-inflammatory effects by modulating inflammatory signaling pathways and inhibiting the release of pro-inflammatory factors such as TNF-α, IL-6, and IL-1β. This dual action endows PBNZ with immense potential for application in the prevention and treatment of diseases[15].
Antioxidant stress effects of PBNZ
As an emerging antioxidant nanozyme, PBNZ demonstrate significant potential in combating oxidative stress. Their key advantage lies in their ability to mimic the activities of multiple natural antioxidant enzymes, achieving a synergistic, multi-pathway antioxidant defense mechanism at the nanoscale. The antioxidant stress effect of PBNZ is primarily attributed to their SOD-like and CAT-like activities[13]. The SOD-like activity of PBNZ enables the dismutation of the chain-reaction-initiating superoxide anion into H2O2 and O2, as shown in the following equation:
This process eliminates ·O2- at its source, converting it into the relatively less toxic H2O2, thereby halting the initiation of the ROS cascade. The CAT-like activity of PBNZ then decomposes the toxic H2O2 into harmless H2O and O2[13], as follows:
This reaction effectively removes the H2O2 generated by the SOD reaction, preventing its conversion into the more destructive ·OH via the Fenton reaction. Consequently, the ROS clearance process mediated by the enzyme-like activities of PBNZ exhibits a synergistic effect: first, SOD-like activity converts ·O2- into H2O2, and then CAT-like activity safely removes H2O2, thus avoiding the accumulation of H2O2 intermediates that may occur with single-enzyme therapy.
The excellent antioxidant stress effects conferred by the CAT-like and SOD-like activities of PBNZ highlight their immense application potential in the treatment of diseases associated with ROS generation. In recent years, researchers have further enhanced their antioxidant efficiency and targeted delivery capabilities through structural modification and functional design of PBNZ. For instance, conjugating PBNZ with molecules possessing targeting recognition functions, such as antibodies or peptides, enables more precise accumulation at disease sites, enhancing ROS scavenging efficacy while minimizing impact on normal tissues[44,45]. PBNZ can also be used in combination with other therapeutic modalities, such as antioxidant drugs or stem cell therapy, to produce synergistic effects, inhibiting oxidative stress responses through multiple pathways and improving the overall efficacy of disease treatment[46] . Furthermore, beyond their role as nanozymes in scavenging ROS, PBNZ can also inhibit oxidative stress through other signaling molecule pathways. In investigating the protective mechanism of PBNZ against sepsis-induced acute lung injury,
Anti-inflammation effects of PBNZ
The anti-inflammatory effect of PBNZ is one of their most significant biological effects. Its mechanism is not a simple, one-way inhibition but rather a complex regulatory network that is multi-target, multi-pathway, and possesses intelligent response characteristics[51]. Inflammation is closely related to oxidative stress[52]. When immune cells are activated, they produce large amounts of reactive oxygen/nitrogen species (ROS/RNS, such as ·O2-, H2O2, ONOO-) as bactericidal mediators. However, excess ROS/RNS can cause self-damage and amplify inflammatory signals. By virtue of their SOD-like and CAT-like activities, PBNZ directly scavenge these ROS/RNS. This not only protects surrounding tissues but, more critically, blocks the activation of inflammatory pathways by ROS as second messengers.
PBNZ can also directly intervene in key inflammatory signaling pathways, thereby exerting anti-inflammatory effects. For example, Zhang et al.[53] recently reported that PBNZ inhibited the inflammatory process of pulpitis by suppressing the NF-κB/NLRP3 pathway, thereby reducing the transmission of downstream inflammatory signals. In a study on using PBNZ to treat Parkinson's disease, Ma et al.[54] found that PBNZ scavenged ROS, which reduced the activation of the NLRP3 inflammasome and caspase-1 in microglia, subsequently downregulating the cleavage of Gasdermin D and the production of inflammatory factors, ultimately inhibiting microglial pyroptosis. In a recent study, our group also found that PBNZ could reduce the release of inflammatory factors (IL-1β, IL-6, and TNF-α) by inhibiting the NF-κB signaling pathway mediated through the activation of the AMP-activated protein kinase (AMPK) signaling pathway. Simultaneously, the O2 produced as an end product of the scavenging of ROS by PBNZ lowers the expression of hypoxia-inducible factor-1α (HIF-1α), which in turn inhibits the expression of its downstream pro-inflammatory factor, iNOS, ultimately exerting a potent anti-inflammatory effect[17].
In addition to exerting anti-inflammatory effects by scavenging ROS/RNS to inhibit oxidative stress and block inflammatory signaling pathways, a growing body of research indicates that PBNZ can also suppress inflammation by regulating macrophage polarization[55-57]. PBNZ can promote the polarization of pro-inflammatory M1 macrophages towards the anti-inflammatory M2 phenotype[17]. This polarization action enables PBNZ to exhibit good therapeutic effects on a variety of inflammatory diseases. For example, the study by Yang et al.[55] showed that PBNZ could attenuate the activation of mitogen-activated protein kinase (MAPK) signaling in M1 macrophages, downregulate the production of pro-inflammatory cytokines (IL-1β and TNF-α), and simultaneously enhance M2 reparative macrophage polarization, thereby effectively treating tendinopathy. Yang et al.[56] also proposed that ultra-small-sized PBNZ could effectively regulate macrophage polarization from pro-inflammatory M1 to anti-inflammatory M2, enhancing the therapeutic outcome of colitis by modulating inflammatory cytokine secretion and restoring intestinal epithelial integrity. Our group's research has also demonstrated that ultra-small-sized PBNZ can repolarize macrophages from a pro-inflammatory M1 to an anti-inflammatory M2 phenotype, thereby effectively combating inflammation in osteoarthritis[57].
Notably, in a recent study using different-sized PBNZ to treat colitis, our group confirmed that the subcellular localization of PBNZ was a critical factor in regulating ROS and macrophage polarization[58]. The results indicated that smaller PBNZ (3 nm) could escape from acidic lysosomes into the neutral cytosol, where they exerted enhanced CAT-like and SOD-like activities, downregulating the phenotype and function of M1 macrophages. In contrast, larger PBNZ (60 and 170 nm) were primarily localized within acidic lysosomes, exhibiting higher POD-like activity and almost no CAT-like activity, which resulted in a diminished capacity for ROS scavenging and M1 macrophage inhibition. This study provides a new perspective, revealing the complex interplay between PBNZ size, lysosomal escape, and M1 macrophage polarization. It also offers a valuable reference for the rational design of prophylactic nanozymes with customized immunomodulatory properties.
THERAPEUTIC EFFECTS OF PBNZ ON IRI
Based on the specific organs or tissues affected, IRI can be categorized into various distinct types. These include myocardial ischemia-reperfusion injury (MIRI), cerebral ischemia-reperfusion injury (CIRI), renal ischemia-reperfusion injury (RIRI), hepatic ischemia-reperfusion injury (HIRI), intestinal ischemia-reperfusion injury, and skin flap ischemia-reperfusion injury, among others. Each type of IRI possesses its own unique pathophysiological characteristics and clinical manifestations. However, oxidative stress and inflammatory responses represent the common pathogenic mechanisms underlying these conditions. Consequently, antioxidant and anti-inflammatory strategies constitute the primary therapeutic approaches for IRI. Due to their favorable SOD- and CAT-mimicking enzymatic activities, PBNZ exhibit excellent capabilities for ROS scavenging and anti-inflammatory effects. Therefore, they represent promising therapeutic candidates for IRI. A growing body of research has reported on the application of PBNZ in treating various types of IRI, including cerebral, myocardial, hepatic, renal, and skin flap IRI, demonstrating promising therapeutic efficacy [Table 1].
PBNZ and its functionalized-modified nanoplatform for the treatment of IRI
| Category of IRI | Types of PBNZ | Model | Delivery method | Biological target | Efficacy | Ref. |
| Cerebral IRI | PBNZ@PVP | MCAOmice | Tail I.V. injection | ROS, inflammation, microglia | ROS scavenging, anti-inflammation, microglia polarization (M1 to M2), neuronal apoptosis inhibition, neurological function recovery | [59] |
| Hollow PBNZ | MCAOmice | In situ injection | RONS, inflammation, MAPK/ERK, PI3K/Akt and NF-κB signal pathway | RONS scavenging, anti-inflammation, anti-apoptosis, neurons protection, negligible side effects | [60] | |
| Sr-substituted Prussian blue (PB)-like nanodrug | CIRI rats | Tail I.V. injection | ROS, inflammation, Ca2+ overload, cGAS-STING signal pathway, mitochondrial DNA | ROS scavenging, anti-inflammation, Ca2+ overload inhibition, mitochondrial function restoration, endoplasmic reticulum stress alleviation, neuronal death reduction | [61] | |
| MPC: melanin (M), Prussian blue (PB) and cerium oxide (C) | CIRI rats | Tail I.V. injection | ROS, inflammation, hypoxia, acidic pH cGAS-STING signal pathway | Sequential targeting from lesions to neuronal mitochondria, ROS scavenging, oxygen regeneration-acid-base balance anti-inflammation, neuronal mitochondria protection, neuronal apoptosis reduction | [16] | |
| A@HPB@THSA | MCAO rats | Tail I.V. injection | ROS, microglia, neutrophil, inflammation, BBB | BBB penetration, Brain targeting, ROS scavenging, microglia polarization (M1 to M2), inhibiting platelet aggregation, suppressing neutrophil infiltration, enhancing neuronal survival | [62] | |
| D@HPB@SPM NPs | MCAO rats | Tail I.V. injection | ROS, inflammation, neutrophil extracellular traps (NETs), BBB | ROS scavenging, Neutrophil hitchhiking, BBB penetration, NETs degradation, alleviating no-flow, neuron protection | [63] | |
| PB-006 @MSC | MCAO mice | Tail I.V. injection | ROS, nNOS-PSD-95 | Targeted drug delivery, reducing immunogenicity, evading immune surveillance, ROS scavenging, neuron protection, | [64] | |
| Myocardial IRI | PBNZ@PSC | MIRI mice | Tail I.V. injection | ROS, inflammation, macrophage, AMPK, NF-κB and HIF-1α signal pathway | ROS scavenging, anti-inflammation, macrophage polarization (M1 to M2), AMPK signaling pathway activation, resistance to hypoxia, vasodilation, reducing MVO and myocardial fibrosis, enhancing cardiac function | [17] |
| DSS/PB @BSP | MIRI mice | Tail I.V. injection | ROS, inflammation, monocytes | Targeting damaged myocardium, ROS scavenging, anti-inflammation, Promoting the polarization of monocytes into M2 type macrophages, improving cardiac function | [65] | |
| Hepatic IRI | PBNZ | HIRI mice | Tail I.V. injection | ROS, inflammation, macrophages | ROS scavenging, anti-inflammation, reducing neutrophil infiltration, macrophages polarization (M1 to M2), reducing hepatocyte apoptosis and tissue damage, restoring liver function | [18] |
| Met@PBN@Neu-CVs | HIRI rats | Tail I.V. injection | ROS, inflammation, Macrophages, mitochondrion | Inflammation targeting, ROS scavenging, anti-inflammation, macrophages polarization (M1 to M2), regulating mitochondrial function and metabolism, reducing liver damage | [66] | |
| PB-MSCs | HIRI mice | Hepatic I.V. injection | ROS, inflammation | ROS scavenging, anti-inflammation, restoring liver function | [67] | |
| Renal IRI | HA-HMPB-Rapa | I/R-induced AKI rats | Tail I.V. injection | ROS, inflammation, mitochondrion | Renal targeting, ROS scavenging, anti-inflammation, inducing mitochondrial autophagy, anti-apoptosis, restoring kidney function | [19] |
| Flap IRI | PBNZ | FIRI mice | In situ injection | ROS, Inflammation, PPAR-γ/NNF-κB, Bax/Bcl-2/caspase-3, and Rip1/Rip3/pMLKL signal pathway | ROS scavenging, anti-inflammation, anti-apoptosis, anti-necroptosis, neovascularization, increasing the survival rate of flaps | [20] |
| Alp@PB-Gel | FIRI mice | In situ injection | ROS, inflammation, NLRP3, CD68, VEGF, CD31 | ROS scavenging, anti-inflammation, promoting angiogenesis, increasing the survival rate of flaps | [68] | |
| UK-PBNZ@PM, | FIRI rats | Tail I.V. injection | ROS, inflammation, thrombus | thrombosis targeting, ROS scavenging, anti-inflammation, reducing thrombosis formation, increasing the survival rate of flaps | [69] |
Cerebral ischemia-reperfusion injury
CIRI refers to the process of further tissue damage that occurs following the restoration of blood flow to brain tissue after a period of ischemia and hypoxia, which can contribute to blood-brain barrier (BBB) disruption, cerebral edema formation, and ultimately, adverse effects on neurological recovery[70]. Clinically, CIRI is commonly observed following thrombolytic or thrombectomy therapy for ischemic stroke, as well as after resuscitation from cardiac arrest. Its occurrence not only worsens the extent of the initial brain injury but also increases the risk of long-term sequelae, potentially manifesting as motor deficits, language dysfunction, and other neurological impairments, thereby negatively impacting patient prognosis[70].
Due to their favorable physicochemical properties and biosafety, PBNZ have demonstrated promising therapeutic potential in CIRI models. Studies have shown that PBNZ can effectively scavenge ROS by mimicking the activities of SOD and CAT, thereby inhibiting oxidative stress and reducing the release of inflammatory factors in CIRI[59]. Furthermore, PBNZ can promote the polarization of microglia from the pro-inflammatory M1 phenotype to the anti-inflammatory M2 phenotype, suppress neuronal apoptosis, and consequently improve neurological recovery following reperfusion in ischemic stroke[59]. The ROS-scavenging efficiency of PBNZ is closely related to its specific surface area. To enhance the therapeutic efficacy of PBNZ against CIRI, Zhang et al.[60] proposed a treatment strategy for ischemic stroke based on hollow Prussian blue nanozymes (HPBNZ). Results showed that HPBNZ exhibited enhanced multi-enzyme-mimicking activities and could effectively scavenge ROS and RONS. Both in vitro and in vivo experiments demonstrated that HPBNZ effectively protected neurons from IRI by mitigating oxidative stress, suppressing inflammatory responses, and inhibiting apoptosis, with minimal side effects.
Innovative modifications of PBNZ to enhance their multifaceted therapeutic effects against CIRI represent a current research hotspot. For instance, Ai's research team constructed a strontium (Sr)-substituted Prussian blue analog nanodrug (SrHCF) for treating CIRI[61]. SrHCF exerts therapeutic effects through multiple mechanisms: scavenging ROS, capturing Fe2+ to transform into PB with stronger antioxidant activity, and releasing Sr2+ to antagonize Ca2+ influx, thereby protecting mitochondrial function and inhibiting neuronal ferroptosis and the inflammatory storm, ultimately significantly alleviating CIRI. Additionally, they innovatively developed a composite nanodrug (MPC) composed of melanin, PB, and CexOy[16]. This system can alleviate CIRI by reprogramming the neuronal microenvironment (NM) through a cascade reaction involving "ROS scavenging-oxygen regeneration-acid-base balance". Both studies represent innovative modifications based on PBNZ, achieving multifaceted therapy for CIRI. The modified materials demonstrated good biocompatibility and safety, providing novel strategies for CIRI treatment.
A critical challenge in CIRI therapy is the difficulty for therapeutic agents to reach the lesion site due to the presence of the BBB. Beyond their inherent antioxidative and anti-inflammatory properties, PBNZ also serve as nanocarriers capable of loading drugs beneficial for CIRI treatment. Through biomimetic design of PBNZ, they can be engineered to effectively cross the BBB and specifically target the lesion site, enabling synergistic targeted therapy for CIRI. For example, Zhang et al.[64] developed a mesenchymal stem cell membrane (MSCm)-based biomimetic co-delivery system (PB-006@MSC) by coating PBNPs loaded with the neuroprotective agent ZL006 with MSC membranes. This system can leverage the chemokine receptors and adhesion proteins on the MSC membrane to achieve active targeting to the ischemic penumbra. It combined the ROS-scavenging capability of PBNPs with the neuroprotective effect of ZL006, significantly reducing cerebral infarct volume, improving neurological function, and lowering mortality [Figure 1A]. Song et al.[62] designed a brain-targeting biomimetic nanozyme (A@HPB@THSA) composed of hollow Prussian blue (HPB) loaded with aspirin (ASA) and modified with T7 peptide-conjugated human serum albumin (THSA), aiming to ameliorate neurovascular unit (NVU) dysfunction after ischemic stroke. This nanozyme achieved targeted enrichment in the ischemic brain region by exploiting transferrin receptor (TfR)-mediated transcytosis on cerebrovascular endothelial cells and increased BBB permeability post-ischemia. The synergistic action of HPB's ROS-scavenging ability and ASA's inhibition of platelet aggregation and neutrophil infiltration protects the NVU [Figure 1B]. Furthermore, their team meticulously designed a biomimetic nanozyme based on a neutrophil-hitchhiking mechanism (D@HPB@SPM NPs)[63]. This nanozyme featured a shell of sialic acid-modified platelet membrane and a core of hollow Prussian blue nanoparticles loaded with deoxyribonuclease I (DNase I). Through specific binding of sialic acid to L-selectin on neutrophil surfaces, it achieves delivery across the BBB to the ischemic brain region
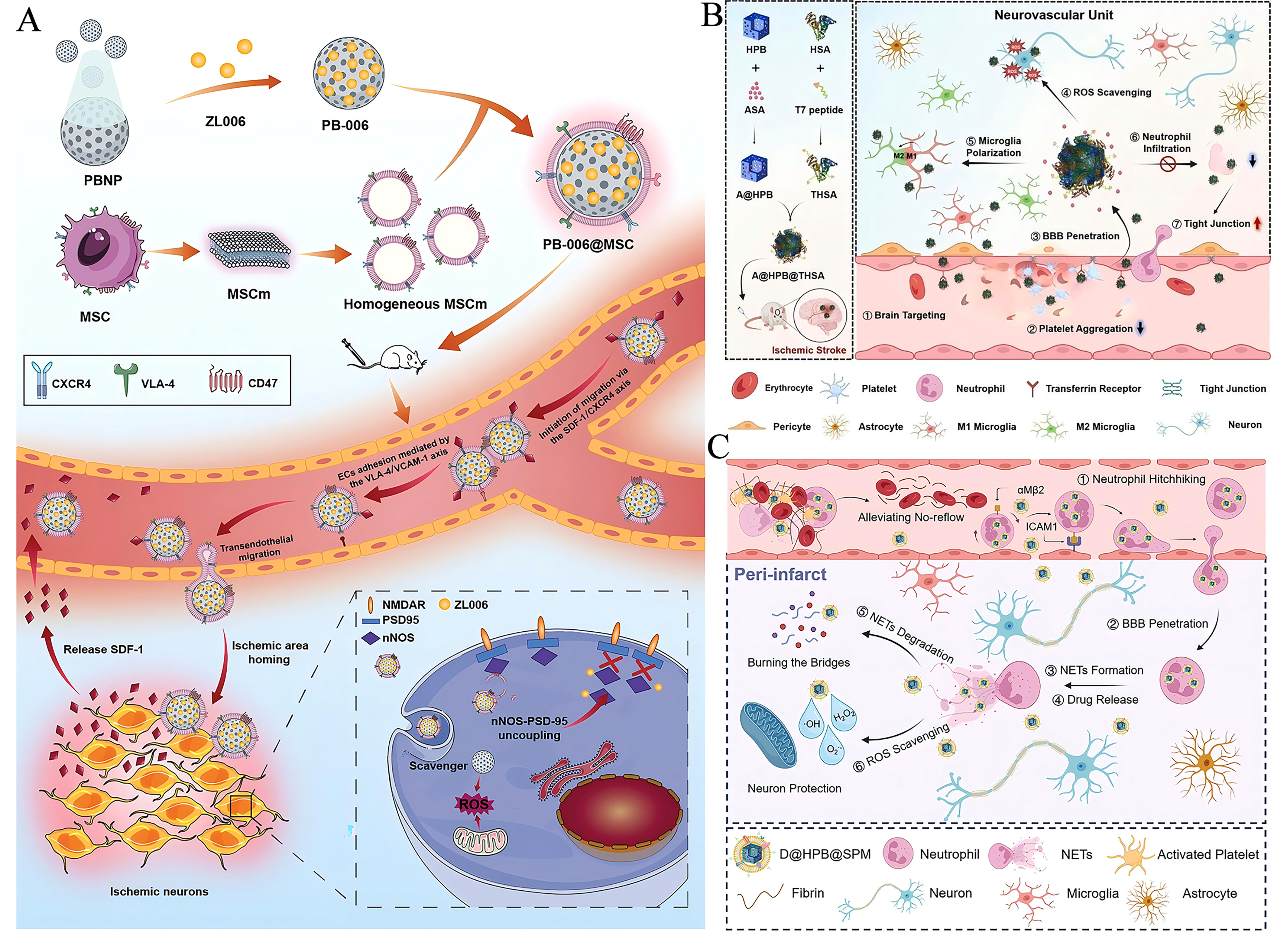
Figure 1. Bionic nano-systems based on PBNZ for the treatment of CIRI. (A) Schematic diagram of the composition of PB-006@MSC and its effect on ischemic brain. This figure is quoted with permission from Zhang et al.[64]. (B) Schematic diagram of the construction of brain-targeted bionic nanoenzyme (A@HPB@THSA) for the treatment of ischemic stroke. This figure is quoted with permission from
Myocardial ischemia-reperfusion injury
MIRI refers to a pathological process in which, despite the restoration of normal perfusion to ischemic myocardium following reperfusion therapies—such as thrombolysis, coronary artery bypass grafting (CABG), or percutaneous coronary intervention (PCI) in patients with acute myocardial infarction (AMI)—the tissue damage progressively worsens. Its principal pathological mechanisms encompass multiple aspects, including oxidative stress burst, calcium overload, mitochondrial dysfunction, and apoptosis. Among these, lipid peroxidation triggered by oxidative stress and the massive release of inflammatory factors are key contributors to the expansion of myocardial cell damage[71]. Consequently, anti-oxidative stress and anti-inflammatory strategies represent the primary therapeutic approaches for MIRI.
PBNZ possesses outstanding antioxidant and anti-inflammatory properties, along with good biocompatibility and safety, making it a potentially effective strategy against MIRI. Recently, our laboratory successfully developed a PBNZ formulation coated with polyglucose sorbitol carboxymethyl ether (PSC), referred to as PBNZ@PSC, and applied it in the treatment of MIRI [Figure 2A][17]. PBNZ@PSC not only exhibits enhanced SOD-like activity due to the polysaccharide nature of its surface coating but also passively targets damaged myocardium via the enhanced permeability and retention (EPR) effect. Both in vitro and in vivo studies have confirmed its excellent biocompatibility, safety, ROS scavenging capacity, and ability to drive macrophage polarization from the M1 to the M2 phenotype, thereby reducing levels of IL-1β, IL-6, and TNF-α to combat inflammation. Thus, PBNZ@PSC can reverse myocardial injury induced by ischemia-reperfusion, reduce the occurrence of coronary microvascular obstruction (MVO), and attenuate myocardial fibrosis, thereby effectively improving cardiac function [Figure 2B]. It is worth emphasizing that this study thoroughly investigated the potential mechanisms underlying PBNZ therapy for MIRI [Figure 2C]. First, after intravenous administration, PBNZ@PSC preferentially targeted damaged myocardium via the EPR effect through compromised vasculature. Subsequently, within cardiomyocytes, PBNZ@PSC utilized its enzyme-like activity to scavenge ROS and generate oxygen, alleviating hypoxia in the infarcted myocardium. On the other hand, it activated the AMPK signaling pathway, promoting endothelial nitric oxide synthase (eNOS) expression to generate nitric oxide (NO), which induces vasodilation, and inhibiting NF-κB signaling pathway to reduce the transcription of inflammatory factors (IL-6, IL-1β, and TNF-α), exerting anti-inflammatory effects. Furthermore, oxygen produced by ROS scavenging and inhibition of the NF-κB pathway by PBNZ@PSC lead to reduced HIF-1α, thereby decreasing the expression of the pro-inflammatory factor inducible iNOS and further contributing to its anti-inflammatory action. This cascade of effects ultimately combats myocardial apoptosis and necrosis, successfully rescuing damaged myocardium. This study provides an in-depth exploration of the detailed anti-inflammatory mechanisms of PBNZ@PSC, offering a solid theoretical foundation for its potential application in treating various inflammatory diseases.
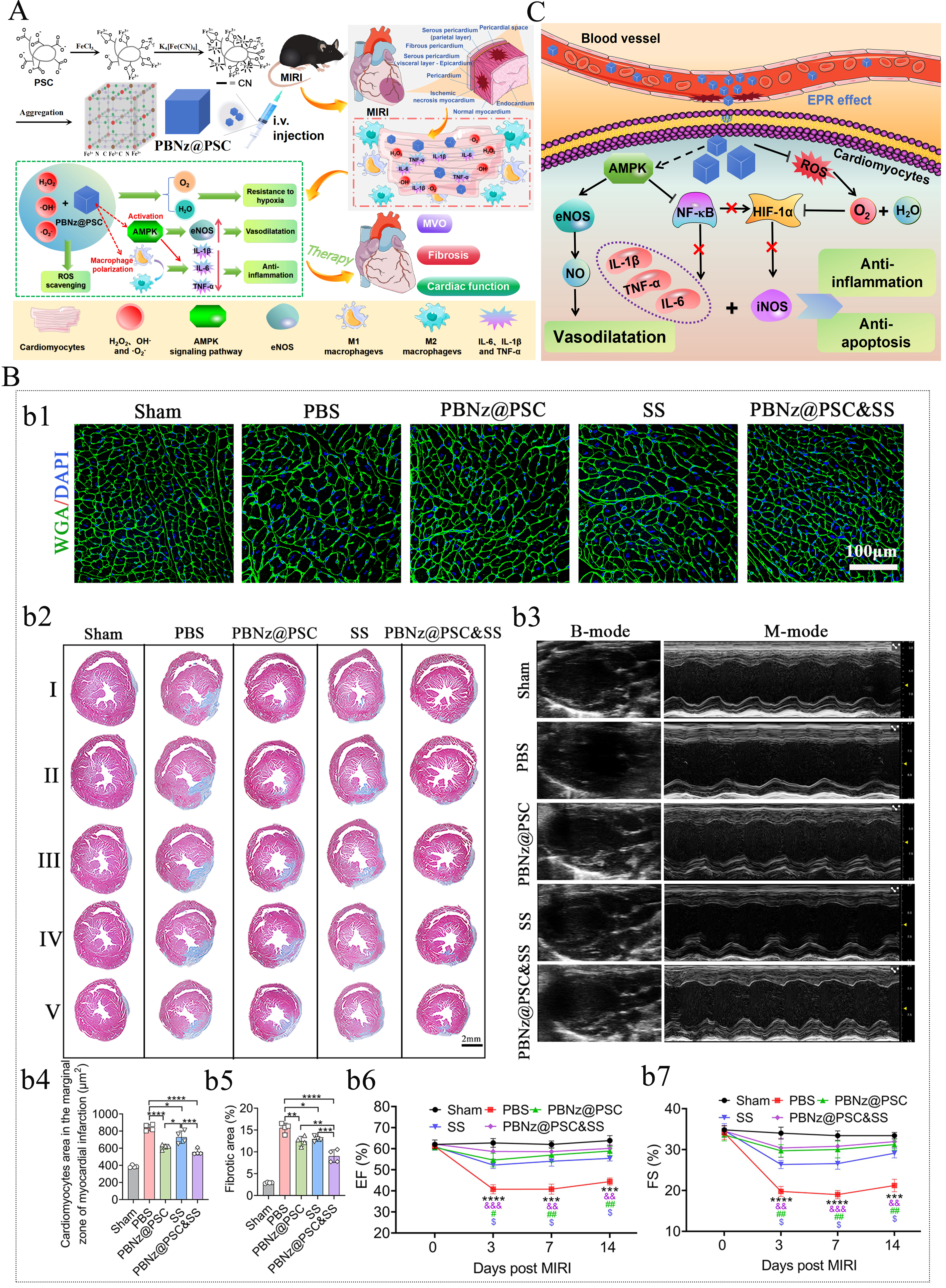
Figure 2. PBNZ@PSC used for the treatment of MIRI. (A) The schematic diagram of the preparation process of PBNZ@PSC and its application in MIRI treatment; (B) The schematic diagram illustrating the possible mechanism of PBNZ@PSC in MIRI treatment; (C) The therapeutic effect of PBNZ@PSC on MIRI. This figure is quoted with permission from Mengmeng Long et al.[17]. Here, * indicates sham vs. PBS, & indicates PBNz@PSC and SS vs. PBS, # indicates PBNz@PSC vs. PBS, and $ indicates SS vs. PBS. Data are presented as mean ± SD, *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001, ****P ≤ 0.0001, &&P ≤ 0.01, &&&P ≤ 0.001, #P ≤ 0.05, ##P ≤ 0.01, and $P ≤ 0.05.
Conventional nanomedicines rely on the EPR effect to target lesion sites. However, this effect is time-sensitive in MIRI treatment, thereby limiting the targeting efficiency of systemic drug delivery. To address this issue, Liu et al.[65] employed hollow Prussian blue nanoparticles as carriers to encapsulate the anti-inflammatory drug betamethasone sodium phosphate (BSP) and further modified them with dextran sulfate sodium (DSS), a targeting ligand for monocyte scavenger receptors, successfully developing DSS/PB@BSP nanoparticles capable of targeting circulating monocytes and reprogramming them into anti-inflammatory M2 macrophages. These nanoparticles can be taken up by monocytes, forming "living cell drug depots" in vivo and leveraging the inflammatory chemotaxis of monocytes to actively transport them to MIRI sites, thereby overcoming the temporal limitations of the EPR effect. Upon reaching the injury site, the nanoparticles scavenged ROS and suppressed inflammation, promoting monocyte polarization into M2 macrophages with anti-inflammatory and reparative functions. This process alleviated inflammatory responses, improved cardiac function, and demonstrated good biosafety, offering a promising non-invasive immunotherapy for MIRI treatment.
Hepatic ischemia-reperfusion injury
HIRI is a pathophysiological process characterized by the paradoxical aggravation of liver tissue damage upon the restoration of blood supply following a period of ischemia and hypoxia. This phenomenon is frequently encountered in clinical scenarios such as liver surgery, traumatic shock, and hepatic inflow occlusion. It constitutes a major cause of post-operative liver dysfunction, primary non-function of the transplanted liver, and even multiple organ failure[72]. Research has indicated that PBNZ demonstrates promising potential for the treatment of HIRI with its antioxidant stress and anti-inflammatory effects[18].
The application of PBNZ for HIRI treatment has already spurred numerous innovative design strategies in this field. For instance, Shen et al.[66] developed a neutrophil membrane-coated PBNZ (Met@PBN@Neu-CVs) for the targeted delivery of metformin (Met) to the ischemia-reperfusion injured liver, enabling precise therapy [Figure 3]. This nanozyme accumulates in the damaged liver via the active targeting function of the neutrophil membrane while simultaneously adsorbing inflammatory factors. Furthermore, it leverages the multi-enzyme activity of PBNZ to synergize with metformin, inhibiting oxidative stress, repairing mitochondrial function, and promoting macrophage polarization toward the anti-inflammatory M2 phenotype. This multi-mechanistic synergistic strategy ultimately alleviates IRI-induced liver damage, providing a comprehensive therapeutic approach for hepatic IRI. Mesenchymal stem cell (MSC) therapy has shown great potential in treating HIRI due to its paracrine and immunomodulatory capacities. However, the low survival rate of transplanted cells in the highly oxidative and stressful microenvironment limits its efficacy. Addressing this, Sahu et al.[67] developed an MSC-based therapy loaded with PBNZ to mitigate HIRI. PBNZ, with its excellent biocompatibility and antioxidant activity, can be efficiently internalized by MSCs without compromising their stemness or differentiation potential. Studies revealed that PB-loaded MSCs (PB-MSCs) exhibited significantly enhanced survival rates under high oxidative stress conditions, along with augmented paracrine and anti-inflammatory functions. Therefore, integrating antioxidant nanozymes into MSCs represents an effective strategy to enhance the efficacy of cell therapy, offering a novel approach for HIRI treatment.
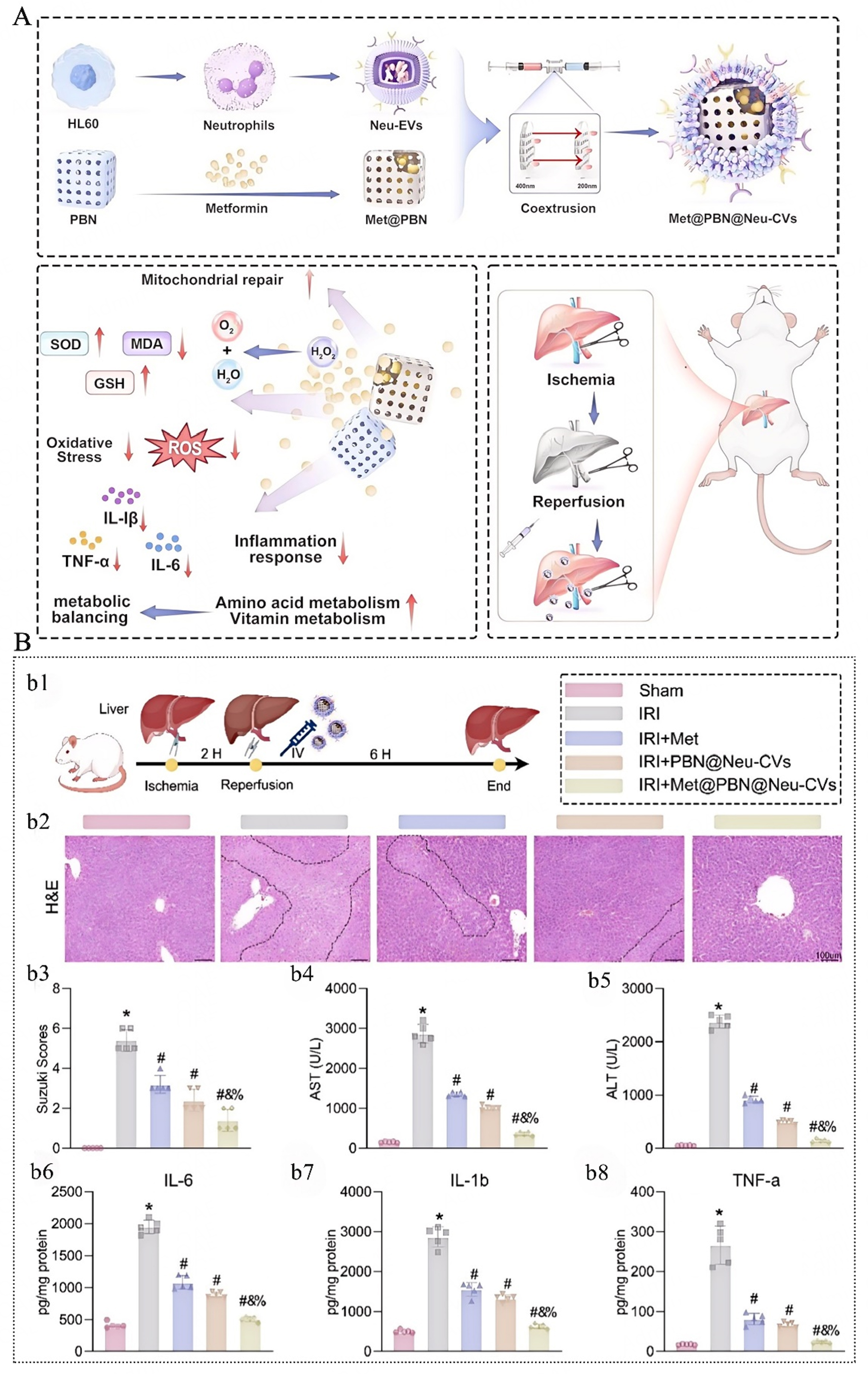
Figure 3. The neutral granulocytes loaded with metformin are coated with PBNZ (Met@PBN@Neu-CVs), which effectively alleviate hepatic ischemia-reperfusion injury (HIRI). (A) Schematic diagram of the synthesis and administration of Met@PBN@Neu-CVs for HIRI treatment; (B) Therapeutic effect of Met@PBN@Neu-CVs on liver IRI in rats. This figure is quoted with permission from Shen et al.[66]. Here, * vs. Sham; # vs. IRI; & vs. IRI + Met; % vs. IRI + PBN@Neu-CVs, P < 0.05, N = 5.
Renal ischemia-reperfusion injury
RIRI frequently occurs following surgeries such as kidney transplantation and partial nephrectomy, leading to damage of renal tubular epithelial cells[73]. This pathological process is a key mechanism underlying acute kidney injury (AKI), potentially causing a sharp decline in renal function and even progression to chronic kidney disease[74]. PBNZ, with their unique anti-inflammatory and antioxidative stress properties, play a crucial role in mitigating tissue damage during ischemia-reperfusion, offering a novel therapeutic opportunity for this challenging medical condition.
Wang et al.[19] developed a targeted drug delivery system for injured kidneys using PBNZ (HA-HMPB-Rapa). In this system, hyaluronic acid (HA) modification enables targeted delivery to CD44-overexpressing injured renal tubular epithelial cells. The hollow mesoporous Prussian blue (HMPB) component, leveraging its nanozyme activity, scavenges ROS, while the loaded rapamycin (Rapa) induces mitophagy. This dual-action strategy effectively clears both ROS and damaged mitochondria. Experimental results demonstrated that this system significantly inhibited oxidative stress, inflammation, and apoptosis in a rat model of I/R-induced AKI. These combined effects synergistically promoted the recovery of renal function, not only reducing the incidence of kidney injury but also substantially lessening its severity. This work represents a novel and effective approach for treating RIRI.
Flap ischemia-reperfusion injury
FIRI, a common complication in plastic and reconstructive surgery, severely impacts flap survival rates and postoperative outcomes[75,76]. Similarly, PBNZ also demonstrated a favorable therapeutic effect on FIRI. As highlighted in a study by Hou et al. [Figure 4][20], PBNZ possesses multiple bioactivities including antioxidant, anti-inflammatory, anti-apoptotic, and anti-necroptotic effects. They significantly improve the microenvironment of I/R-injured flaps by scavenging ROS and modulating key pathways: the PPAR-γ/NF-κB inflammatory pathway, the Bax/Bcl-2/caspase-3 apoptotic pathway, and the Rip1/Rip3/pMLKL necroptotic pathway. This intervention increased the flap survival rate from 37.21% ± 8.21% to 79.61% ± 7.5%, effectively enhancing flap viability and providing a new strategy for clinical flap management.
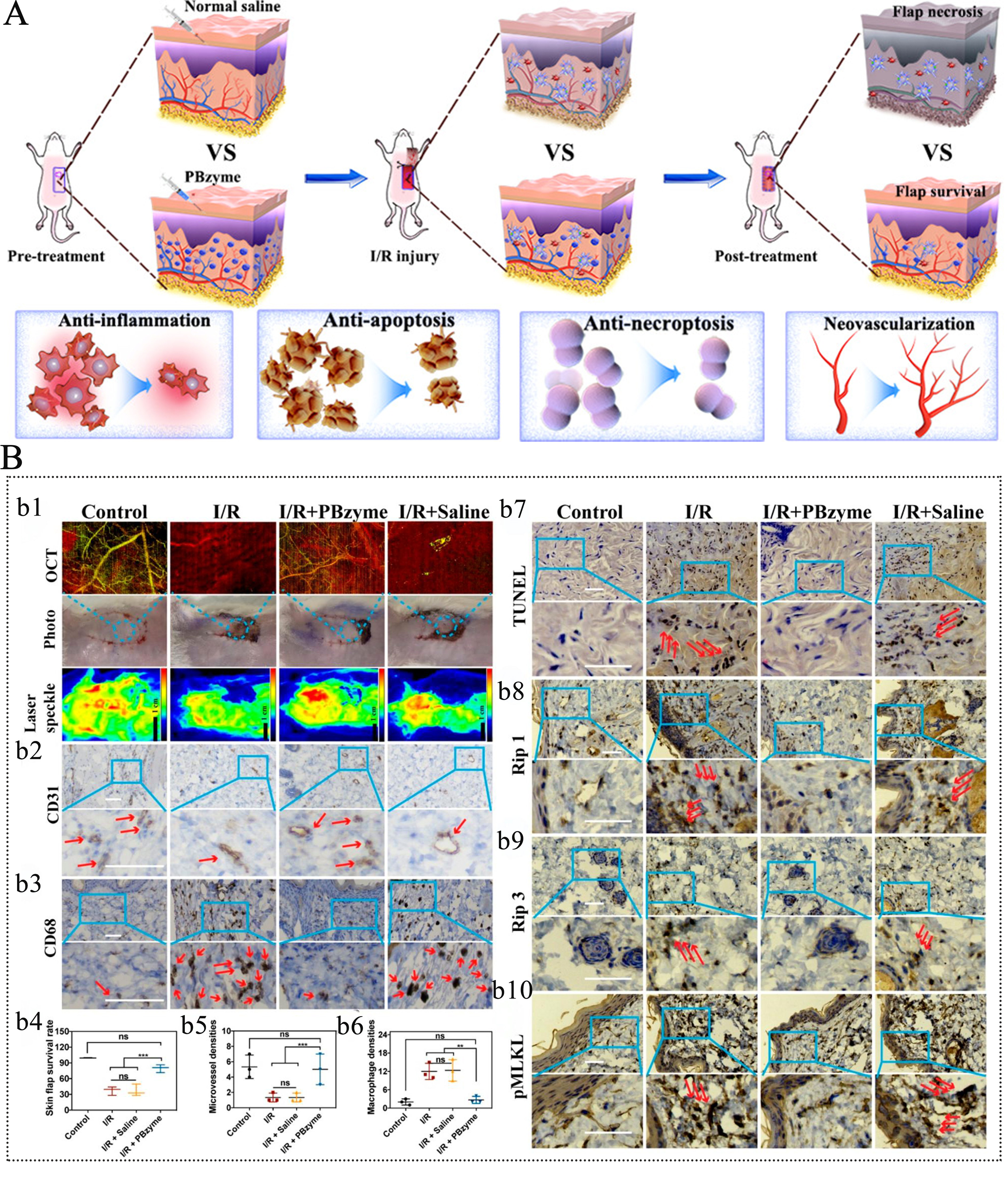
Figure 4. PBNZ improves the survival rate of skin flaps in ischemia-reperfusion injury. (A) Schematic diagram of the application of PBNZ in the treatment of flap ischemia-reperfusion injury; (B) Therapeutic effect of PBNZ on flap ischemia-reperfusion injury. This figure is quoted with permission from Hou et al.[20]. Here, **P < 0.05; ***P < 0.001; ns: not significantly different; N = 3.
Furthermore, functional modification of PBNZ, combined with their utility as drug carriers to co-deliver therapeutic agents, can form a targeted synergistic treatment system, which is expected to significantly improve the therapeutic efficacy against FIRI. For instance, Liu's research team developed a porcine acellular dermal matrix gel loaded with alprostadil (Alp) and PBNPs, termed Alp@PB-Gel, for FIRI treatment[68]. In this composite, the extracellular matrix (ECM) hydrogel provided a regenerative microenvironment and served as a drug reservoir, PBNPs contributed ROS-scavenging and anti-inflammatory properties, and alprostadil (Alp) acted as a vasodilator. Thus, the Alp@PB-Gel integrated the advantages of all three components, exhibiting both cell affinity and plasticity. And, the photothermal effect of PBNPs allowed for controlled drug release, achieving synergy between short-term vasodilation and long-term regenerative repair. In vitro experiments confirmed its ability to scavenge ROS and promote the proliferation of vascular endothelial cells and angiogenesis. In vivo studies demonstrated its efficacy in alleviating FIRI by downregulating NLRP3 and CD68 to inhibit inflammation and apoptosis, and upregulating VEGF and CD31 to promote vascular regeneration, significantly improving flap survival with good biocompatibility and clinical potential.
In order to address both IRI and the thrombotic microenvironment in flap transplantation, Yang et al.[69] proposed an ischemia time-based sequential treatment strategy. They constructed a clinical-grade biomimetic vesicle, UK-PBNZ@PM, for FIRI therapy. This system integrates the thrombolytic function of urokinase (UK), the anti-inflammatory and antioxidant capabilities of PBNZ, and the targeted delivery ability of platelet membranes (PM). Through the dual mechanisms of thrombolysis and IRI prevention, it reduced thrombus formation and cleared the intravascular inflammatory microenvironment, ultimately enhancing the survival rate of ischemic flaps. These innovative PBNZ-based therapeutic strategies are not only operationally feasible but also target multiple pathological links in FIRI, providing strong experimental evidence and potential treatment protocols for clinically improving the success rate of flap transplantation.
CONCLUSION AND PERSPECTIVES
In recent years, with the rapid development of nanomaterial preparation technologies and in-depth biomedical research, PBNZ have shown great application potential in IRI therapy. With unique multi-enzyme activity-mimicking properties, PBNZ can modulate key pathophysiological processes like oxidative stress and inflammatory responses in complex biological environments, offering a new perspective for IRI treatment. In addition, as a drug carrier, PBNZ can achieve targeted drug delivery and controlled release through surface modification or functional design, thereby further enhancing the precision and safety of treatment. Meanwhile, its excellent biocompatibility and biodegradability provide a safeguard against potential toxic side effects associated with long-term use, offering a novel approach to address the issues of low delivery efficiency and significant side effects in conventional IRI therapies. From basic research to pre-clinical exploration, PBNZ have had significant therapeutic effects in IRI models of various tissues and organs, such as the brain, myocardium, liver, kidneys, and skin flaps. However, many challenges need to be addressed for the translation of PBNZ from laboratory research to clinical application.
Firstly, the long-term biosafety of PBNZ must be highly prioritized. Although current research shows good biocompatibility, issues like the in vivo metabolic pathways of nanomaterials, potential accumulation effects, and possible immunological responses need more in-depth and long-term evaluation. A comprehensive understanding of these potential risks is essential for the safety and reliability of PBNZ in clinical applications.
Secondly, the targeted delivery efficiency of PBNZ and their accumulation capacity at lesion sites need further improvement. Achieving more specific delivery of PBNZ to injured tissues and reducing impact on normal tissues via more precise surface modification techniques or intelligent responsive designs is crucial for improving therapeutic efficacy and minimizing toxic side effects. This requires not only innovations in material design but also the integration of advanced biomedical technologies to achieve higher targeting specificity and lower side effects.
Furthermore, research into the mechanism of action of PBNZ requires further deepening and refinement. In particular, their interaction with complex in vivo biomolecular networks and the observed differences in their effects across various types of IRI models demand more systematic, comprehensive, and in-depth investigations to elucidate.
Moreover, large-scale preparation processes and strict quality control standards for PBNZ should be established and improved. Scientific production procedures and quality management systems are needed to ensure batch-to-batch consistency and reliability for clinical use. Meanwhile, systematic pharmacokinetic and pharmacodynamic studies on PBNZ are required to provide a scientific basis for clinical dosing regimens. These studies will help understand its in vivo distribution, metabolism, and mechanism of action, thus optimizing clinical application strategies.
Looking to the future, with the continuous integration and development of interdisciplinary research, and through collaborative innovation and joint efforts across multiple fields such as materials science, biology, and medicine, we have reason to believe that the performance of PBNZ will be further optimized, and their application scope will continue to expand. This, in turn, will bring new breakthroughs and hope for the clinical treatment of IRI.
DECLARATIONS
Authors’ contributions
Writing-original draft and visualization: Long M
Writing review & editing: Han X
Writing-review & editing and supervision: Sheng J, Gu N
Availability of data and materials
Not applicable.
AI and AI-assisted tools statement
Not applicable.
Financial support and sponsorship
This work was supported by the National Key Research and Development Program of China (No. 2024YFF0508600), National Natural Science Foundation of China (No. 61821002, 32301189) and the Natural Science Foundation of Jiangsu Province (Grant No. BK20222002).
Conflicts of interest
All authors declared that there are no conflicts of interest.
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Copyright
© The Author(s) 2026.
REFERENCES
1. Zhang M, Liu Q, Meng H, et al. Ischemia-reperfusion injury: molecular mechanisms and therapeutic targets. Sig Transduct Target Ther. 2024;9:12.
2. Soares ROS, Losada DM, Jordani MC, Évora P, Castro-E-Silva O. Ischemia/Reperfusion injury revisited: an overview of the latest pharmacological strategies. Int J Mol Sci. 2019;20:5034.
3. Nieuwenhuijs-Moeke GJ, Pischke SE, Berger SP, et al. Ischemia and reperfusion injury in kidney transplantation: relevant mechanisms in injury and repair. J Clin Med. 2020;9:253.
4. Goncharov RG, Sharapov MG. Ischemia-reperfusion injury: molecular mechanisms of pathogenesis and methods of their correction. Mol Biol. 2023;57:1143-64.
5. Wu M, Yiang G, Liao W, et al. Current mechanistic concepts in ischemia and reperfusion injury. Cell Physiol Biochem. 2018;46:1650-67.
6. Xin W, Qin Y, Lei P, Zhang J, Yang X, Wang Z. From cerebral ischemia towards myocardial, renal, and hepatic ischemia: exosomal miRNAs as a general concept of intercellular communication in ischemia-reperfusion injury. Mol Ther Nucl Acids. 2022;29:900-22.
7. Deng J, Liu Q, Ye L, et al. The Janus face of mitophagy in myocardial ischemia/reperfusion injury and recovery. Biomed Pharmacother. 2024;173:116337.
8. He S, Liu CY, Ren CH, Zhao H, Zhang XX. Immunological landscape of retinal ischemia-reperfusion injury: insights into resident and peripheral immune cell responses. Aging Dis. 2025;16:115-36.
9. Jurcau A, Simion A. Neuroinflammation in cerebral ischemia and ischemia/reperfusion injuries: from pathophysiology to therapeutic strategies. Int J Mol Sci. 2021;23:14.
10. Pefanis A, Ierino FL, Murphy JM, Cowan PJ. Regulated necrosis in kidney ischemia-reperfusion injury. Kidney Int. 2019;96:291-301.
11. Lu K, Zhu X, Li Y, Gu N. Progress in the preparation of Prussian blue-based nanomaterials for biomedical applications. J Mater Chem B. 2023;11:5272-300.
12. Qin Z, Li Y, Gu N. Progress in applications of Prussian blue nanoparticles in biomedicine. Adv Healthc Mater. 2018;7:1800347.
13. Zhang W, Hu S, Yin J, et al. Prussian blue nanoparticles as multienzyme mimetics and reactive oxygen species scavengers. J Am Chem Soc. 2016;138:5860-5.
14. He H, Long M, Duan Y, Gu N. Prussian blue nanozymes: progress, challenges, and opportunities. Nanoscale. 2023;15:12818-39.
15. Wang Y, Liang Z, Liang Z, Lv W, Chen M, Zhao Y. Advancements of Prussian blue-based nanoplatforms in biomedical fields: progress and perspectives. J Controlled Release. 2022;351:752-78.
16. Wang S, Li R, Shi X, et al. Sequential targeted nanomedicine MPC treats cerebral ischemia-reperfusion injury via neuronal microenvironment reprogramming. Chem Eng J. 2025;524:169755.
17. Long M, Wang L, Kang L, et al. Prussian blue nanozyme featuring enhanced superoxide dismutase-like activity for myocardial ischemia reperfusion injury treatment. ACS Nano. 2025;19:4561-81.
18. Huang Y, Xu Q, Zhang J, et al. Prussian blue scavenger ameliorates hepatic ischemia-reperfusion injury by inhibiting inflammation and reducing oxidative stress. Front Immunol. 2022;13:891351.
19. Wang Y, Fu H, Huang B, et al. Hollow mesoporous Prussian blue nanoenzymes as Rapamycin carrier for targeted treatment of ischemia/reperfusion-induced acute kidney injury through pro-mitophagy and oxidative stress alleviation. Chem Eng J. 2024;497:155684.
20. Hou R, Lu T, Gao W, et al. Prussian blue nanozyme promotes the survival rate of skin flaps by maintaining a normal microenvironment. ACS Nano. 2022;16:9559-71.
21. Peserico D, Stranieri C, Garbin U, et al. Ezetimibe prevents ischemia/reperfusion-induced oxidative stress and up-regulates Nrf2/ARE and UPR signaling pathways. Antioxidants. 2020;9:349.
22. Dugbartey GJ. Cellular and molecular mechanisms of cell damage and cell death in ischemia-reperfusion injury in organ transplantation. Mol Biol Rep. 2024;51:473.
23. Güler MC, Tanyeli A, Ekinci Akdemir FN, et al. An overview of ischemia-reperfusion injury: review on oxidative stress and inflammatory response. Eurasian J Med. 2023;54:S62-5.
24. Eltzschig HK, Eckle T. Ischemia and reperfusion—from mechanism to translation. Nat Med. 2011;17:1391-401.
25. Malkov MI, Lee CT, Taylor CT. Regulation of the hypoxia-inducible factor (HIF) by pro-inflammatory cytokines. Cells. 2021;10:2340.
26. Halladin NL. Oxidative and inflammatory biomarkers of ischemia and reperfusion injuries. Dan Med J. 2015;62:1-22.
27. Wu L, Xiong X, Wu X, et al. Targeting oxidative stress and inflammation to prevent ischemia-reperfusion injury. Front Mol Neurosci. 2020;13:28.
28. Peterson JM, Smith TA, Rock EP, Magnani JL. Selectins in biology and human disease: opportunity in E-selectin antagonism. Cureus. 2024;16:1-24.
29. Lv W, Liu Y, Li S, et al. Nano drug delivery system based on natural cells and derivatives for ischemic stroke treatment. Chin Med J. 2025;138:1945-60.
30. Dri E, Lampas E, Lazaros G, et al. Inflammatory mediators of endothelial dysfunction. Life. 2023;13:1420.
31. Puhl S, Steffens S. Neutrophils in post-myocardial infarction inflammation: damage vs. resolution? Front Cardiovasc Med. 2019;6:25.
32. Schofield ZV, Woodruff TM, Halai R, Wu MC, Cooper MA. Neutrophils - A key component of ischemia-reperfusion injury. Shock. 2013;40:463-70.
33. Duan W, Wang X, Ma Y, et al. Therapeutic strategies targeting the NLRP3‑mediated inflammatory response and pyroptosis in cerebral ischemia/reperfusion injury (Review). Mol Med Rep. 2024;29:46.
34. Gorsuch WB, Chrysanthou E, Schwaeble WJ, Stahl GL. The complement system in ischemia-reperfusion injuries. Immunobiology. 2012;217:1026-33.
35. De Oliveira THC, Gonçalves GKN. Complement system: a key player in hepatic ischemia-reperfusion injury. Biochem Biophys Res Commun. 2025;783:152580.
36. Hou Z, Brenner JS. Developing targeted antioxidant nanomedicines for ischemic penumbra: Novel strategies in treating brain ischemia-reperfusion injury. Redox Biol. 2024;73:103185.
37. Agbadua OG, Kúsz N, Berkecz R, Gáti T, Tóth G, Hunyadi A. Oxidized resveratrol metabolites as potent antioxidants and xanthine oxidase inhibitors. Antioxidants. 2022;11:1832.
38. Hou L, Zhao M, Huang C, He Q, Zhang L, Zhang J. Alternative oxidase gene induced by nitric oxide is involved in the regulation of ROS and enhances the resistance of Pleurotus ostreatus to heat stress. Microb Cell Fact. 2021;20:137.
39. He P, Talukder MAH, Gao F. Oxidative stress and microvessel barrier dysfunction. Front Physiol. 2020;11:472.
40. Neri M, Fineschi V, Paolo M, et al. Cardiac oxidative stress and inflammatory cytokines response after myocardial infarction. Curr Vasc Pharmacol. 2015;13:26-36.
41. Allowitz K, Taylor J, Harames K, Yoo J, Baloch O, Ramana KV. Oxidative stress-mediated lipid peroxidation-derived lipid aldehydes in the pathophysiology of neurodegenerative diseases. Curr Neuropharmacol. 2025;23:671-85.
42. Ambroz A, Klema J, Rossnerova A, et al. Associations of environmental pollution with pro-oxidant, antioxidant and inflammatory markers in pregnant mothers and newborns. Front Toxicol. 2025;7:1572486.
43. Rocco E, Grimaldi MC, Maino A, et al. Advances and challenges in biomarkers use for coronary microvascular dysfunction: from bench to clinical practice. J Clin Med. 2022;11:2055.
44. He H, Han Q, Wang S, et al. Design of a multifunctional nanozyme for resolving the proinflammatory plaque microenvironment and attenuating atherosclerosis. ACS Nano. 2023;17:14555-71.
45. Bai H, Wang T, Kong F, et al. CXCR4 and CD44 dual-targeted Prussian blue nanosystem with daunorubicin loaded for acute myeloid leukemia therapy. Chem Eng J. 2021;405:126891.
46. Ding X, Hu Y, Feng X, et al. Enhanced blood‐brain barrier penetrability of BACE1 SiRNA‐loaded prussian blue nanocomplexes for Alzheimer's disease synergy therapy. Exploration. 2025;5:e20230178.
47. Li X, Qiu C, Yang F, Li X, Fang Y, Yang Y. Protective effects of Prussian blue nanozyme against sepsis-induced acute lung injury by activating HO-1. Eur J Pharmacol. 2024;968:176354.
48. Wei W, Wang Y, Sun G, Chen Z, Zhang S. Prussian blue nanoparticles mitigate inflammatory osteolysis by reducing oxidative stress and enhancing endogenous antioxidant systems. Colloids Surf B Biointerfaces. 2025;254:114806.
49. Bai H, Kong F, Feng K, et al. Prussian blue nanozymes prevent anthracycline-induced liver injury by attenuating oxidative stress and regulating inflammation. ACS Appl Mater Interfaces. 2021;13:42382-95.
50. Feng Q, Xu H, Pan X, et al. Antioxidation and anti-inflammatory activity of Prussian blue nanozymes to alleviate acetaminophen-induced acute liver injury. ACS Appl Nano Mater. 2023;6:8468-81.
51. Li D, Liu M, Li W, et al. Synthesis of Prussian blue nanoparticles and their antibacterial, antiinflammation and antitumor applications. Pharmaceuticals. 2022;15:769.
52. Ramos-González E, Bitzer-Quintero O, Ortiz G, Hernández-Cruz J, Ramírez-Jirano L. Relationship between inflammation and oxidative stress and its effect on multiple sclerosis. Neurologia. 2024;39:292-301.
53. Zhang L, Ning X, Cui J, et al. Thermosensitive injectable in situ forming hydrogel incorporating anti-oxidative stress and anti-inflammatory nanoparticles for vital pulp therapy of pulpitis. Mater Today Biol. 2025;31:101482.
54. Ma X, Hao J, Wu J, Li Y, Cai X, Zheng Y. Prussian blue nanozyme as a pyroptosis inhibitor alleviates neurodegeneration. Adv Mater. 2022;34:2106723.
55. Yang J, Chen J, Liu Y, et al. Multifunctional Prussian blue nanozymes ameliorate tendinopathy via modulating tissue homeostasis. Mater Today Biol. 2025;34:102187.
56. Yang G, Li Q, Meng J, Dong H, Qin Z, Mou Y. Ultra-small Prussian blue nanodots scavenge reactive oxidative species and balance M1/M2 macrophages to mitigate colitis inflammation. Nanoscale. 2025;17:15402-12.
57. Qin Z, Li X, Wang P, et al. Ultrasmall Prussian blue nanozyme attenuates osteoarthritis by scavenging reactive oxygen species and regulating macrophage phenotype. Nano Lett. 2024;24:11697-705.
58. Ding H, Long M, Wu Y, et al. Subcellular distribution of Prussian blue nanozymes dictates enzymatic activity and macrophage polarization for effective colitis therapy. Biomaterials. 2026;325:123622.
59. Liu J, Sun J, Song Y, et al. Prussian blue nanozyme treatment of ischemic brain injury via reducing oxidative stress inhibits inflammation, suppresses apoptosis, and promotes neurological recovery. ACS Chem Neurosci. 2023;14:1535-46.
60. Zhang K, Tu M, Gao W, et al. Hollow Prussian blue nanozymes drive neuroprotection against ischemic stroke via attenuating oxidative stress, counteracting inflammation, and suppressing cell apoptosis. Nano Lett. 2019;19:2812-23.
61. Xiong T, Wang S, Shi X, et al. Biomimetic dynamic nanomedicine SrHCF: lattice self-reconstruction based on Fe2+ response to achieve synergistic “ferrous chelation-antioxidation-calcium antagonism-inflammation inhibition” quadruple treatment of CIRI. Bioact Mater. 2026;55:23-41.
62. Song Y, Wang L, Peng T, et al. Brain-targeting biomimetic nanozyme enhances neuroprotection in ischemic stroke by remodeling the neurovascular unit. J Controlled Release. 2025;382:113750.
63. Song J, Yang G, Song Y, et al. Neutrophil hitchhiking biomimetic nanozymes prime neuroprotective effects of ischemic stroke in a tailored “burning the bridges” manner. Adv Funct Mater. 2024;34:2315275.
64. Zhang Q, Li S, Chen H, et al. Reduction of oxidative stress and excitotoxicity by mesenchymal stem cell biomimetic co-delivery system for cerebral ischemia-reperfusion injury treatment. Small. 2024;20:2401045.
65. Liu Y, Zhou M, Xu M, et al. Reprogramming monocytes into M2 macrophages as living drug depots to enhance treatment of myocardial ischemia-reperfusion injury. J Controlled Release. 2024;374:639-52.
66. Shen P, Huang K, Zhang X, Qin M, Wang X, Fan Z. Neutrophil-mimicking Prussian blue nanozymes for synergistic targeted immuno-metabolic therapy of hepatic ischemia-reperfusion injury. J Nanobiotechnol. 2025;23:736.
67. Sahu A, Jeon J, Lee MS, Yang HS, Tae G. Nanozyme impregnated mesenchymal stem cells for hepatic ischemia-reperfusion injury alleviation. ACS Appl Mater Interfaces. 2021;13:25649-62.
68. Xiong X, Yin C, Tong A, et al. Dermal extracellular matrix gelatin delivering Prussian blue nanoparticles to relieve skin flap ischemia. Int J Biol Macromol. 2024;267:131361.
69. Yang L, Liu Y, Tao C, et al. Bionic nanovesicles sequentially treat flaps with different durations of ischemia by thrombolysis and prevention of ischemia-reperfusion injury. Mater Today Bio. 2025;31:101529.
70. Yang M, Liu B, Chen B, Shen Y, Liu G. Cerebral ischemia-reperfusion injury: mechanisms and promising therapies. Front Pharmacol. 2025;16:1613464.
71. Du B, Fu Q, Yang Q, et al. Different types of cell death and their interactions in myocardial ischemia-reperfusion injury. Cell Death Discov. 2025;11:87.
72. George J, Lu Y, Tsuchishima M, Tsutsumi M. Cellular and molecular mechanisms of hepatic ischemia-reperfusion injury: The role of oxidative stress and therapeutic approaches. Redox Biol. 2024;75:103258.
73. Chen Y, Yang L. Cellular senescence in renal ischemia-reperfusion injury. Chin Med J. 2025;138:1794-806.
74. Li C, Yu Y, Zhu S, et al. The emerging role of regulated cell death in ischemia and reperfusion-induced acute kidney injury: current evidence and future perspectives. Cell Death Discov. 2024;10:216.
75. Liu S, Xiong X, Chen L, et al. Targeting programmed cell death in flap ischemia/reperfusion injury. Biomolecules. 2025;15:911.
Cite This Article
How to Cite
Download Citation
Export Citation File:
Type of Import
Tips on Downloading Citation
Citation Manager File Format
Type of Import
Direct Import: When the Direct Import option is selected (the default state), a dialogue box will give you the option to Save or Open the downloaded citation data. Choosing Open will either launch your citation manager or give you a choice of applications with which to use the metadata. The Save option saves the file locally for later use.
Indirect Import: When the Indirect Import option is selected, the metadata is displayed and may be copied and pasted as needed.
About This Article
Copyright

Data & Comments
Data
0











Comments
Comments must be written in English. Spam, offensive content, impersonation, and private information will not be permitted. If any comment is reported and identified as inappropriate content by OAE staff, the comment will be removed without notice. If you have any queries or need any help, please contact us at support@oaepublish.com.