

How photoenzymatic radical reactions are integrated into microbial biosynthesis
 0
0 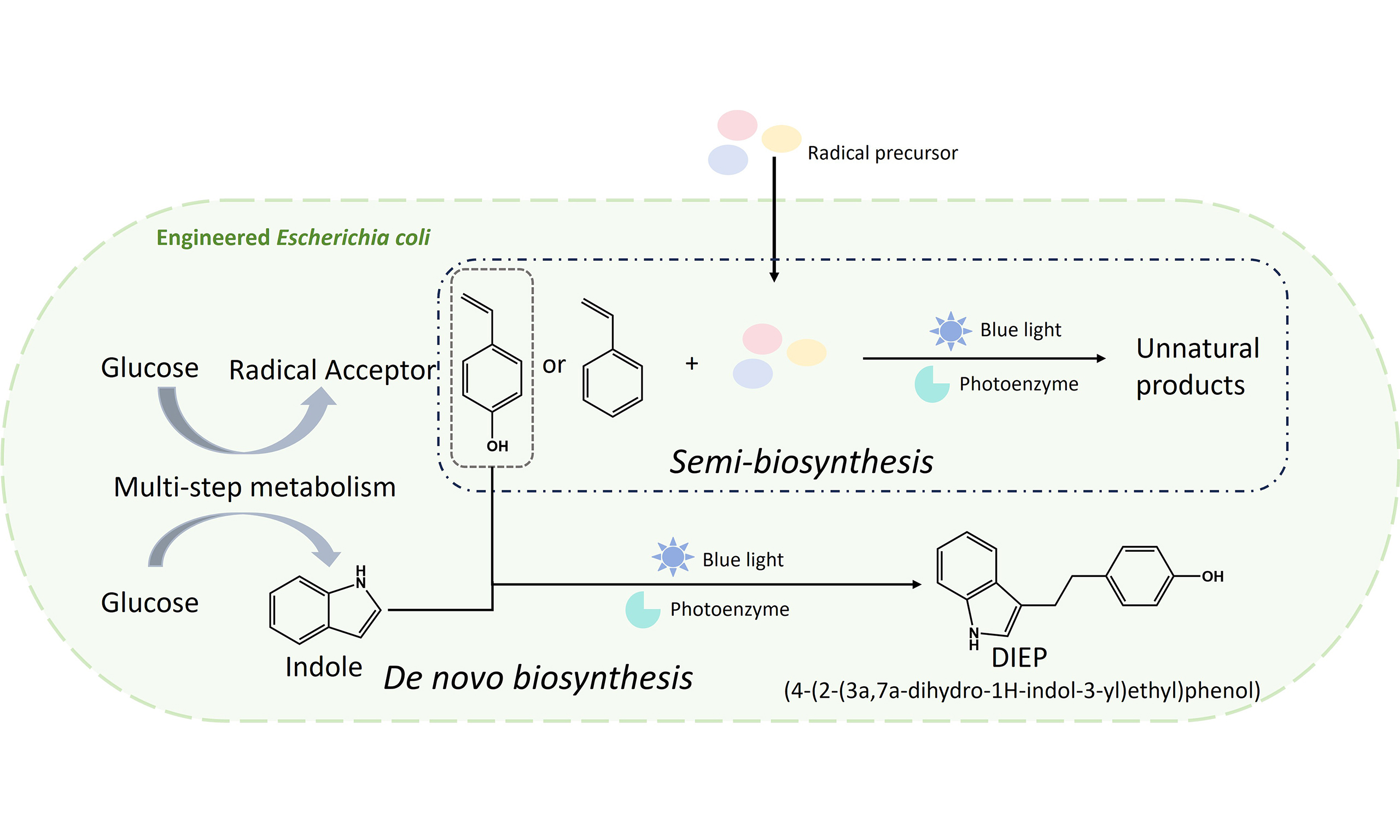
Biosynthesis offers unparalleled precision for constructing complex molecular architectures, yet its chemical scope remains fundamentally limited by the reactivity of natural enzymes[1]. Extending biosynthesis beyond nature’s enzymatic repertoire has therefore motivated the development of new strategies within synthetic biology[2-4]. In this context, photoenzymatic catalysis has emerged as a promising approach, coupling enzymatic selectivity with light-driven radical activation to access transformations that are otherwise difficult to achieve[5-7]. However, most photoenzymatic reactions have been confined to in vitro systems or tightly constrained whole-cell formats, where high enzyme loading, dependence on exogenous cofactors and limited operational stability hinder translation toward scalable production[5,6]. In contrast, Yuan et al.[1] establish an in vivo photobiosynthesis strategy that integrates photoenzymatic radical chemistry into microbial metabolism[8,9], enabling the continuous production of non-natural molecules within living systems. This work highlights the potential for scalable and sustainable biosynthesis in microbial systems.
To establish this in vivo photobiosynthesis platform, photoenzymatic radical reactivity was systematically integrated with intracellular metabolic processes. In vitro screening of flavin-dependent ene-reductases and their variants under blue-light irradiation enabled the identification of an engineered variant of Saccharomyces cerevisiae old yellow enzyme 3 with high activity toward radical fluorination. As shown in previous studies[10], visible light enables access to the flavin semiquinone state of the bound cofactor, allowing for single-electron transfer and controlled radical C–C bond formation within enzyme active sites. Attention was then directed toward substrate availability within the cell. A biosynthetic pathway for 4-vinylphenol production was reconstructed in Escherichia coli, together with genetic modifications that enhance precursor flux from central metabolism. This design enabled intracellular generation of the olefinic radical acceptor [Figure 1]. Integration of the photoenzyme with this substrate-producing system resulted in light-dependent formation of fluorinated products. In contrast, strains lacking either photoenzyme expression or endogenous substrate production exhibited little or no conversion. Subsequent optimisation focused on coordinating photoenzyme activity with cellular reducing capacity and substrate flux. Modulation of reduced nicotinamide adenine dinucleotide phosphate (NADPH)-regeneration pathways, promoter strengths and plasmid copy numbers led to stepwise improvements in product titres. Notably, intracellular biosynthesis of the radical acceptor proved more effective than exogenous substrate supplementation, underscoring the importance of localised metabolite generation for sustaining photoenzymatic radical reactions in vivo.
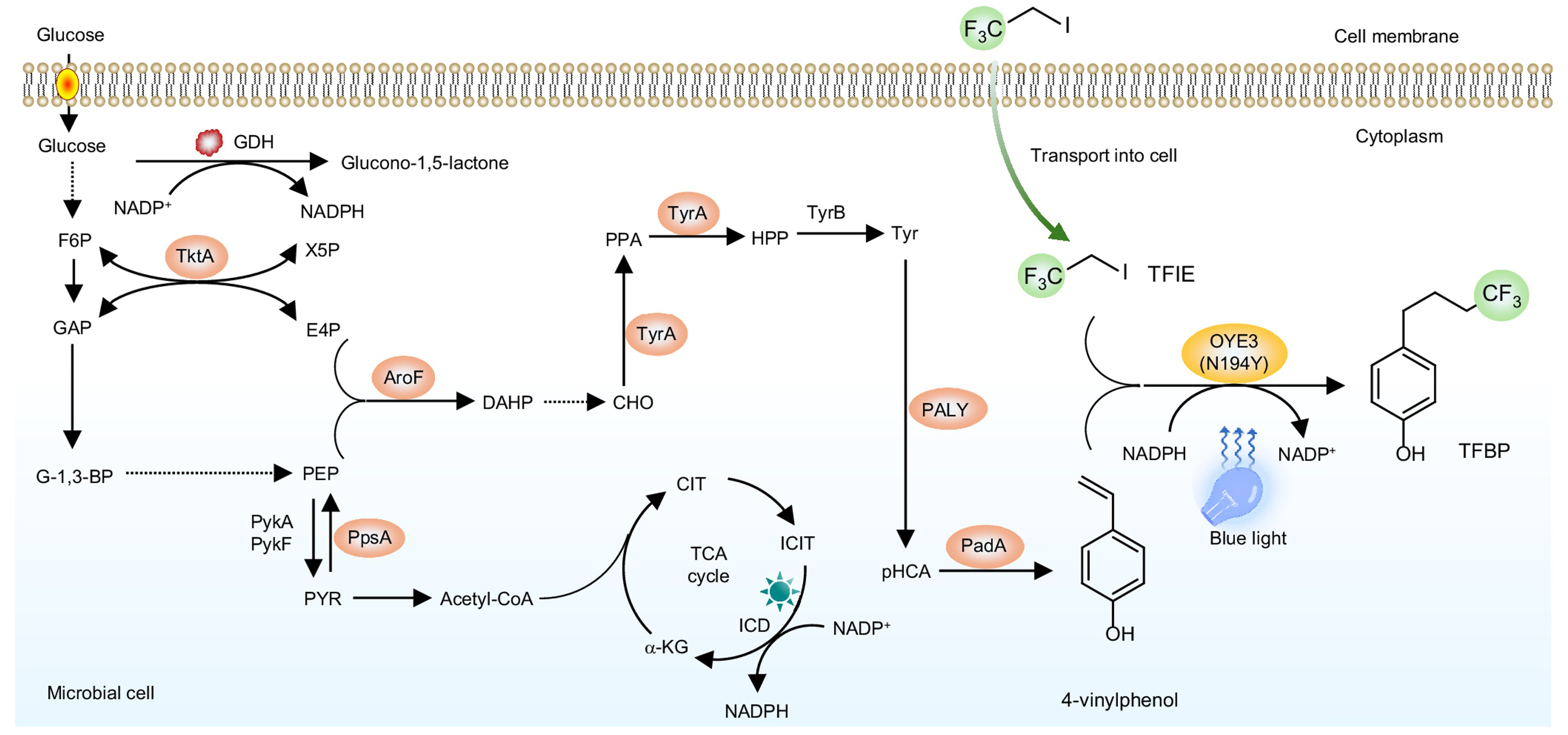
Figure 1. Semi-synthetic photobiosynthesis in engineered Escherichia coli combining endogenous olefin production with externally supplied radical precursors. Adapted with permission from[1], Copyright 2026, Springer Nature. AroF: 3-Deoxy-D-arabino-heptulosonate-7-phosphate (DAHP) synthetase; CHO: chorismate; CIT: citrate; E4P: erythrose 4-phosphate; F6P: fructose 6-phosphate; GAP: glyceraldehyde 3-phosphate; HPP: 4-hydroxyphenylacetone; ICD: isocitrate dehydrogenase; ICIT: isocitrate; α-KG: α-ketoglutarate; G-1,3-BP: glycerate-1,3-bisphosphate; NADPH: reduced nicotinamide adenine dinucleotide phosphate; Tyr: tyrosine; TyrA: tyrosine synthase; TyrB: tyrosine aminotransferase; TktA: transketolase; X5P: xylulose 5-phosphate; PALY: phenylalanine ammonia lyase (from R. toruloides); PadA: decarboxylase (from L. plantarum); PEP: phosphoenolpyruvate; pHCA: p-hydroxy-cinnamic acid; PPA: prephenate; PpsA: phosphoenolpyruvate synthase; PykA/F: pyruvate kinase A/F; PYR: pyruvate; TCA: tricarboxylic acid; TFIE: trifluoroiodoethane; TFBP: 4-(4,4,4-trifluorobutyl) phenol; OYE3N194Y: old yellow enzyme 3 (from Saccharomyces cerevisiae) with substitution of asparagine for tyrosine at amino acid residue 194.
The in vivo photobiosynthesis platform further demonstrated its generality by accommodating both new substrates and diverse classes of photoenzymatic reactions. In addition to 4-vinylphenol, intracellular biosynthesis of styrene enabled light-driven functionalisation of a second, structurally distinct olefin within the same microbial framework. Across these endogenously supplied substrates, the platform supported multiple photoenzymatic radical transformations, including hydroalkylation, hydroamination and hydrosulfonylation with diverse radical precursors, giving access to a range of non-natural products [Figure 2]. Importantly, asymmetric photoenzymatic reactions proceeded in vivo with stereochemical outcomes comparable to those observed in vitro, indicating that integration of radical photochemistry in a cellular environment does not compromise catalytic control. The platform also demonstrated compatibility with conditions relevant to scale-up. By decoupling aerobic substrate formation from anaerobic, light-driven radical catalysis, oxygen-sensitive photoenzymatic reactions became operable within fermentation systems. Implementation under controlled bioreactor conditions enabled sustained photoreactivity and increased product accumulation during fed-batch operation. Together, these features establish the platform as a broadly adaptable framework, capable of extending photoenzymatic radical chemistry across substrates, reaction types and different scales of implementation within living microbial systems.
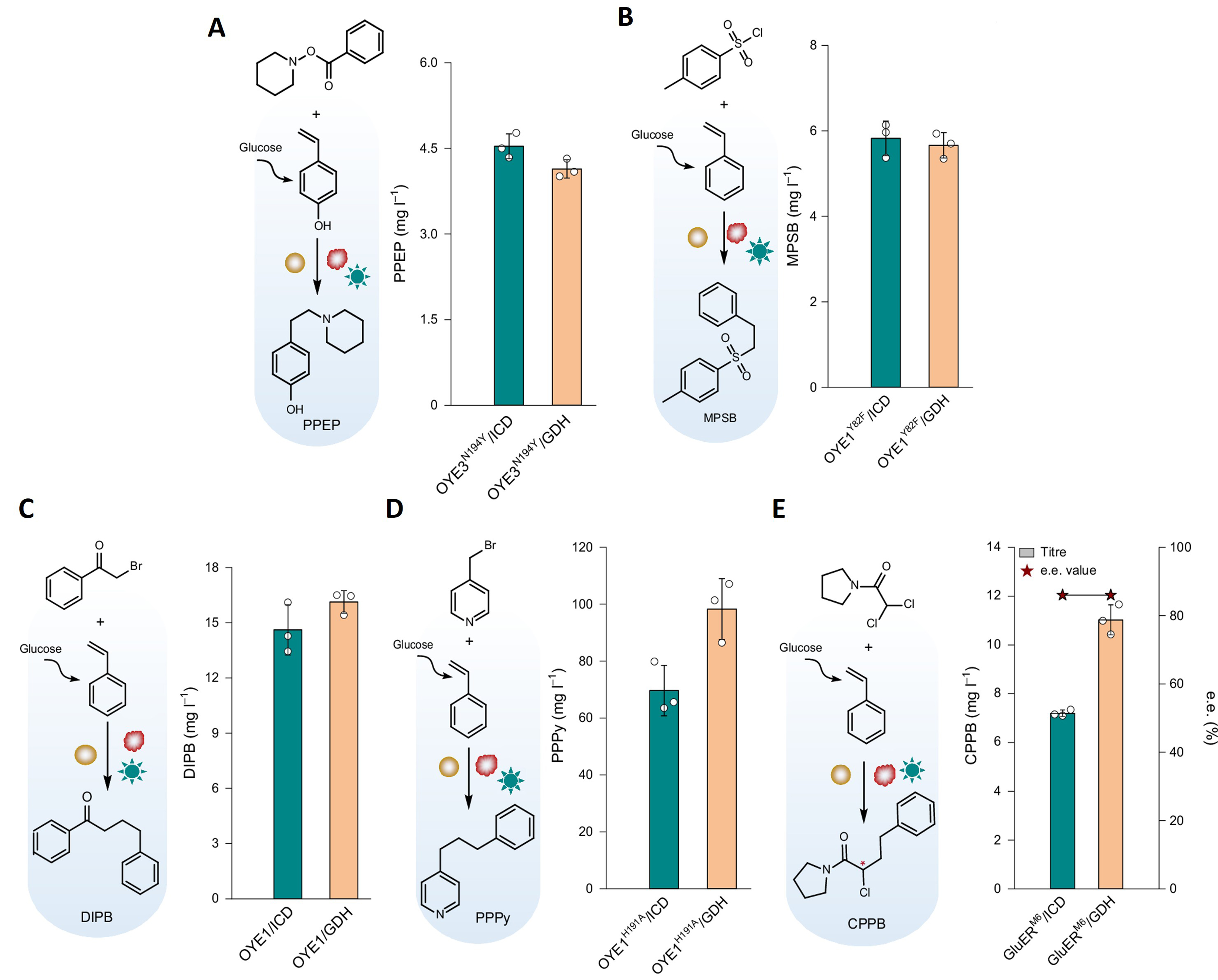
Figure 2. Scope of photoenzymatic radical transformations enabled by semi-integrated photobiosynthesis: (A) hydroamination, (B) hydrosulfonylation, and (C-E) hydroalkylation. Adapted with permission from[1], Copyright 2026, Springer Nature. ICD: Isocitrate dehydrogenase; GDH: glucose dehydrogenase; OYE3: ene-reductases (from Saccharomyces cerevisiae); OYE1: ene-reductases (from Saccharomyces pastorianus); GluER: ene-reductase (from Gluconobacter oxydans); PPEP: 4-(2-(piperidin-1-yl)ethyl)phenol; DIPB: 1,4-diphenylbutan-1-one; MPSB: 1-methyl-4-(phenethylsulfonyl)benzene; PPPy: 4-(3-phenylpropyl) pyridine; CPPB: 2-chloro-4-phenyl-1-(pyrrolidin-1-yl)butan-1-one.
To further advance the platform from semi-synthetic operation toward full pathway integration, photoenzymatic reactions were incorporated into a single microbial host in which the radical precursor, olefin substrate and photoenzyme were all produced intracellularly [Figure 3A]. In this configuration, indole biosynthesis was coupled with endogenous generation of 4-vinylphenol and photoenzyme expression, enabling light-driven hydroarylation without the need for external feeding of key reactants. This integrated system consequently enables the biosynthesis of 4-(2-(3a,7a-dihydro-1H-indol-3-yl)ethyl)phenol (DIEP), a structurally complex phenol-indole analogue, which has been preliminarily shown to exhibit broad-spectrum antimicrobial activity[1]. Rather than operating as an add-on transformation supported by supplied substrates, photoenzymatic catalysis proceeds within the context of cellular metabolism, illustrating a transition from partially integrated to fully integrated photobiosynthesis.
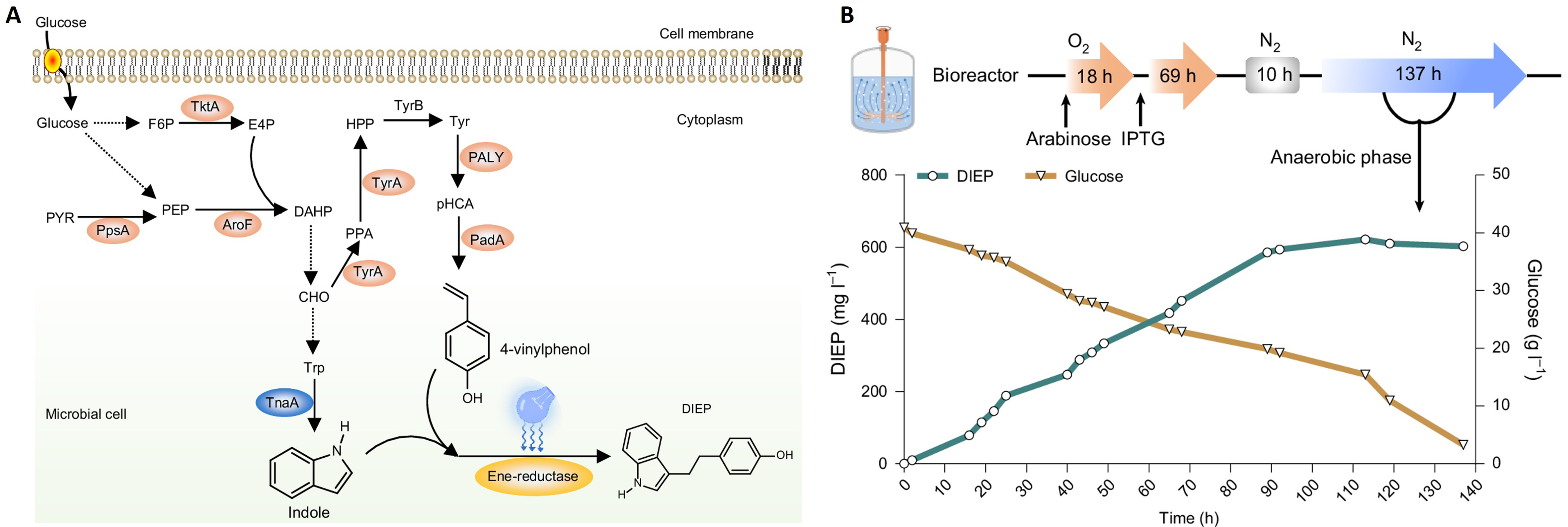
Figure 3. Fully integrated photobiosynthesis and fermentation-enabled production of DIEP. (A) Intracellular generation of the olefin substrate, radical precursor, and photoenzyme enables de novo photobiosynthesis; (B) Fed-batch fermentation supports sustained, light-driven production through separation of metabolic and catalytic processes. Adapted with permission from[1], Copyright 2026, Springer Nature. DIEP: 4-(2-(3a,7a-dihydro-1H-indol-3-yl)ethyl)phenol; F6P: fructose 6-phosphate; E4P: erythrose 4-phosphate; TktA: transketolase; PYR: pyruvate; PpsA: phosphoenolpyruvate synthase; PEP: phosphoenolpyruvate; AroF: 3-deoxy-D-arabino-heptulosonate-7-phosphate (DAHP) synthetase; CHO: chorismate; Trp: tryptophan; TnaA: tryptophanase; Tyr: tyrosine; TyrA: tyrosine synthase; TyrB: tyrosine aminotransferase; PPA: prephenate; HPP: 4-hydroxyphenylacetone; PALY: phenylalanine ammonia lyase (from R. toruloides); PadA: decarboxylase (from L. plantarum); pHCA: p-hydroxy-cinnamic acid.
The resulting integrated system was further stabilised and intensified through coordinated genetic and process-level optimisation. Adjustment of expression levels and cofactor availability improved reaction efficiency, while protein engineering enhanced the activity of the photoenzyme in vivo. Implementation in fed-batch fermentation enabled the separation of aerobic metabolite accumulation from anaerobic, light-driven catalysis, supporting sustained product formation. Sequential regulation of parallel biosynthetic modules further reduced pathway interference, underscoring the role of regulatory balance in maintaining efficient photobiosynthesis under productive conditions [Figure 3B]. These efforts demonstrate how increasing levels of integration and control enable photoenzymatic reactions to be maintained and amplified in microbial hosts.
Overall, this work establishes a microbial photobiosynthesis platform in which photoenzymatic catalysis is directly integrated into cellular metabolism, enabling light-driven radical transformations to operate in vivo rather than as isolated in vitro reactions. By combining flavin mononucleotide (FMN)-dependent ene-reductases with engineered biosynthetic pathways in Escherichia coli[1], the system supports both semi-biosynthesis mediated by externally supplied radical precursors and fully integrated de novo biosynthesis in which the photoenzyme, radical precursor and substrate are produced intracellularly. This strategy enables sustained, fermentation-based production of a range of non-natural molecules and culminates in the complete biosynthesis of a complex, bioactive compound under light irradiation. Collectively, these results demonstrate that photoenzymatic reactions can be systematically coupled to microbial production pathways and maintained within a living, renewable reaction environment.
Despite these advances, the current strategy still faces several limitations that define the practical boundaries of in vivo photobiosynthesis. Reaction performance remains constrained by the complexity of coordinating light-driven catalysis with cellular metabolism, highlighting challenges in achieving robust, general and well-controlled reaction conditions in living systems. Addressing these limitations will require further improvements in system stability, substrate scope and overall process controllability. Nonetheless, by demonstrating that light-driven enzymatic transformations can be stabilised, intensified and scaled under fermentation conditions, this work lays a foundation for the development of sustainable molecular manufacturing strategies. It creates opportunities for the biosynthetic production of high-value pharmaceuticals, bioactive molecules and structurally complex compounds through photoenzymatic transformations, positioning living cells as viable platforms for executing chemistries beyond the reach of natural enzyme catalysis[1].
DECLARATIONS
Authors’ contributions
Drafted the manuscript: Zheng, Y.
Revised and rewrote sections of the manuscript: Hu, J.; Li, Z.; Abraham, B.; Obry, A.
Availability of data and materials
Not applicable.
AI and AI-assisted tools statement
Not applicable.
Financial support and sponsorship
None.
Conflicts of interest
Hu, J. is a Section Editor of the journal Chemical Synthesis. Hu, J. was not involved in any steps of editorial processing, notably including reviewers’ selection, manuscript handling, or decision-making. The other authors declare that there are no conflicts of interest.
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Copyright
© The Author(s) 2026.
REFERENCES
1. Yuan, Y.; Li, M.; Harrison, W.; Zhang, Z.; Zhao, H. Harnessing photoenzymatic reactions for unnatural biosynthesis in microorganisms. Nat. Catal. 2026, 9, 62-72.
2. Yang, Y.; Arnold, F. H. Navigating the unnatural reaction space: directed evolution of heme proteins for selective carbene and nitrene transfer. Acc. Chem. Res. 2021, 54, 1209-25.
3. Kissman, E. N.; Sosa, M. B.; Millar, D. C.; Koleski, E. J.; Thevasundaram, K.; Chang, M. C. Y. Expanding chemistry through in vitro and in vivo biocatalysis. Nature 2024, 631, 37-48.
4. Brouwer, B.; Della-Felice, F.; Illies, J. H.; Iglesias-Moncayo, E.; Roelfes, G.; Drienovská, I. Noncanonical amino acids: bringing new-to-nature functionalities to biocatalysis. Chem. Rev. 2024, 124, 10877-923.
5. Emmanuel, M. A.; Bender, S. G.; Bilodeau, C.; et al. Photobiocatalytic strategies for organic synthesis. Chem. Rev. 2023, 123, 5459-520.
6. Yu, J.; Chen, B.; Huang, X. Single-electron oxidation triggered by visible-light-excited enzymes for asymmetric biocatalysis. Angew. Chem. Int. Ed. Engl. 2025, 64, e202419262.
7. Peng, Y.; Chen, Z.; Xu, J.; Wu, Q. Recent advances in photobiocatalysis for selective organic synthesis. Org. Process. Res. Dev. 2022, 26, 1900-13.
8. Huang, J.; Liu, Z.; Bloomer, B. J.; et al. Unnatural biosynthesis by an engineered microorganism with heterologously expressed natural enzymes and an artificial metalloenzyme. Nat. Chem. 2021, 13, 1186-91.
9. Huang, J.; Quest, A.; Cruz-Morales, P.; et al. Complete integration of carbene-transfer chemistry into biosynthesis. Nature 2023, 617, 403-8.
Cite This Article
How to Cite
Download Citation
Export Citation File:
Type of Import
Tips on Downloading Citation
Citation Manager File Format
Type of Import
Direct Import: When the Direct Import option is selected (the default state), a dialogue box will give you the option to Save or Open the downloaded citation data. Choosing Open will either launch your citation manager or give you a choice of applications with which to use the metadata. The Save option saves the file locally for later use.
Indirect Import: When the Indirect Import option is selected, the metadata is displayed and may be copied and pasted as needed.
About This Article
Copyright

Data & Comments
Data
0













Comments
Comments must be written in English. Spam, offensive content, impersonation, and private information will not be permitted. If any comment is reported and identified as inappropriate content by OAE staff, the comment will be removed without notice. If you have any queries or need any help, please contact us at support@oaepublish.com.